 2017, Vol. 38
2017, Vol. 38


2. 宝鸡文理学院地理与环境学院, 宝鸡 721013
2. College of Geography and Environmental Engineering, Baoji University of Arts and Sciences, Baoji 721013, China
土壤呼吸是一个复杂的生态学过程, 其主要包括根系呼吸、微生物呼吸以及地表凋落物呼吸[1], 且在全球尺度上, 每年因地上和地下凋落物矿化和分解而释放到大气中的CO2浓度占到总土壤呼吸的比例高达70%以上[2].地表凋落物作为联系地上和地下部分的重要枢纽, 在林地生态系统碳循环中起着重要作用[3, 4], 地表凋落物一方面可以通过腐殖化以土壤有机碳的形式存储在土壤中[4], 另一方面可以通过土壤微生物的矿化和分解以土壤呼吸的形式释放到大气中[3].因此, 地表凋落物呼吸的变化特征与土壤中的碳储量和气候变暖趋势(大气CO2浓度增加)密切相关.
在林地生态系统中, 地表凋落物呼吸是土壤呼吸的重要组成部分, 而地表凋落物呼吸占土壤呼吸的比例大约为10%~40%[2, 5~9], 且这一比例在水热条件较好的热带和亚热带地区高达30%以上[10~12], 在水热条件适中的的温带地区维持在20%左右[8, 13, 14], 在寒温带地区由于较低的大气/土壤温度而小于10%[5, 6].土壤微环境(土壤温度和水分)是影响凋落物呼吸的重要因素[9, 15], 且土壤温度和土壤水分对凋落物呼吸的影响分别利用指数和抛物线关系模型进行拟合[9, 16, 17].有研究表明, 凋落物呼吸温度敏感性变化于1.29~10.6之间[7, 16, 18, 19], 且凋落物呼吸温度敏感性的时空变异性与土壤温度[18]、土壤水分[20]等因素有关.在旱地生态系统中, 除了土壤温度和水分以外, 降雨的剧烈波动性不可避免地影响到土壤水分的有效性, 而土壤水分有效性将进一步影响归还到土壤中的地表凋落物量[21, 22], 而底物的数量也是影响凋落物呼吸温度敏感性变异的主要因素之一[18].因此, 笔者推测除了土壤温度和水分以外, 降水导致的地表凋落物量变化也是影响凋落物呼吸温度敏感性的重要因素之一.
在林地生态系统中, 针对土壤呼吸/土壤呼吸温度敏感性[23~25]、根系和微生物呼吸/根系和微生物呼吸温度敏感性[17, 18, 26]、甚至地表凋落物呼吸/地表凋落物呼吸贡献率[6, 8, 9, 13, 16]的变化特征及其影响因素均有大量的研究.虽然也有相关的研究探讨凋落物呼吸温度敏感性的变化特征[7, 16~20], 但是其内在的驱动机制仍然很匮乏.基于此, 本研究在黄土高原一个恢复了30年的典型人工刺槐林内, 通过地表凋落物控制试验(对照处理、去除地表凋落物处理、倍增地表凋落物处理)利用5年连续的实测资料(土壤呼吸、土壤温度、土壤水分、地表凋落物), 研究凋落物呼吸温度敏感性的变化特征及其影响因素, 以期为气候变化条件下地区的土壤呼吸与气候反馈机制提供数据支撑和理论依据.
1 材料与方法 1.1 地点描述王东沟小流域(35°13′N, 107°40′E; 海拔1 095 m)位于陕西省长武县, 为中国科学院长武黄土高原农业生态试验站(长武农田生态试验站)所在地.流域属于黄土高原南部典型高塬沟壑区, 其地貌类型由塬面、沟坡以及沟道三部分组成, 且这三部分约各占土地面积的1/3, 沟壑密度为2.78条·km-2.塬面海拔1 220 m, 流域内从塬面到沟底的最大高差为280 m.塬面主要用于粮食的生产, 而沟坡由于陡峻的地形以及不合理的人为活动极易遭受水土流失, 其土壤侵蚀模数高达50 [t·(hm2·a)-1], 而严重的水土流失已经极大地威胁到该地区的粮食安全、地表水质量, 改变了地区的水文循环模式.
流域属半干旱湿润性季风气候. 1984~2011年间年均降水量为586 mm, 其中最高年份为954 mm, 最低年份为296 mm, 6~8月降水量占年总量的60%左右.年平均气温9.4℃, 大于10℃积温为3 029℃, 年日照时数为2 230 h, 日照率为51%, 年辐射总量为484 kJ·cm-2, 无霜期171 d.表层土壤(0~20 cm)有机碳6.50 g·kg-1, 全氮0.62 g·kg-1, 碱解氮37.0 mg·kg-1, 速效磷3.0 g·kg-1, 速效钾129.3 mg·kg-1, CaCO310.5%, pH 8.4, 黏粒含量(<0.002 mm) 14%, 田间持水量22.4%, 土壤凋萎含水量9.0%[14, 16].
1.2 植被描述在黄土区, 人工刺槐林(Robinia pseudoacacia L.)于20世纪70年代被广泛种植(种植密度为1 213株·hm-2), 目的是为了遏制严重的水土流失和改善其恶劣的生态环境.在该林地生态系统中(0.68 hm2), 平均树高(H)约为(6.8±1.6) m, 胸径(DBH)为(6.4±2.6) cm, 植被覆盖度为55%.林下灌丛以茅莓(Rubus parvifolius L.)为主, 其树高和覆盖度分别为(62.8±11.8) cm和55%, 林下草本植物以白羊草(Bothriochloa ischaemum (L.) Keng)为主, 其草高为(42.5±11.9) cm, 郁闭度为75%[14, 16].
1.3 试验设计在2008年3月, 于王东沟小流域内的典型沟坡系统上, 随机选择生长相对均一的人工刺槐林为研究对象, 试验设置了3种地表凋落物控制措施:对照处理、去除地表凋落物处理、倍增地表凋落物处理[8, 16, 27], 并于每个地表凋落物控制措施下建立5个1.5 m×1.5 m的永久试验小区.在对照处理中(LCK), 地上部凋落物被允许正常地输入小区内部; 在去除地表凋落物处理中(LR), 地上部凋落物被网眼大小为1 mm2的尼龙网从小区中移除; 而在倍增地表凋落物处理中(LA), 人为定期(月初)地将从去除地表凋落物处理下收集的地表凋落物量均匀的撒在倍增地表凋落物处理下, 其目的是实现该处理下的地表凋落物量翻倍.同时为了减少风对不同地表凋落物控制处理下凋落物的保持尽可能不产生或者少产生影响, 将试验小区放在林子的中间.
在整个试验期间(2009~2013), 于林地上方布置了5个尼龙网(面积为1 m2; 网眼大小为1 mm2)以此来收集地上部凋落物量.地表凋落物量每季度收集一次, 收集后的地表凋落物水洗后置于烘箱中在70℃的恒温下烘72 h, 待地表凋落物质量保持恒重后称重[g·(m2·a)-1][28].
1.4 土壤呼吸、土壤温度和水分的测量土壤呼吸的测定利用闭路式土壤碳通量系统, 仪器型号为LI-8100 (直径20 cm, LI-COR, licon, NE, USA).为了避免PVC管安装过程中对土壤造成的扰动, 继而引起短期土壤呼吸的剧烈波动.因此, 在初次测定土壤呼吸前的24 h安装PVC管(直径20 cm, 高12 cm), 并使PVC管出露地表 2 cm, 安装好的PVC管永久留在试验小区内, 直至试验结束.土壤呼吸测定前去除PVC管里面的一切可见的地上部动植物活体, 每个PVC管上连续测定两次, 两次测量之间的时间间隔为30 s, 这两次土壤呼吸值之间的变异控制在15%之内.每个PVC管上的测量时间为150 s, 其中包括30 s的前期预处理、30 s的后期预处理以及90 s的观测期.试验期间(2009-03~2013-11) 每天在09:00~11:00之间进行土壤呼吸的测定, 此时间段内的测定值可代表一天的平均值[29, 30], 平均大概每15 d测定一次.冬季(12月、1月和2月)由于气候寒冷, 土壤温度和水分波动较小, 植物和土壤中的微生物均处于休眠状态, 因此冬季不进行土壤呼吸的测定.土壤呼吸在试验期间依次测量了13、12、15、19和13次.
在测定土壤呼吸的同时测定土壤温度(每个PVC管3次重复)和水分(每个PVC管4次重复), 土壤温度和水分的测定在距离PVC管10 cm处进行.距地表 5 cm处的土壤温度利用数字显示温度计(仪器型号:JM624测温仪; 产地:中国; 精度:±0.2 FS±0.1℃)测定, 0~5 cm处的土壤水分利用便携式水分仪(仪器型号:ML3-HH2;产地:英国; 精度:±1% vol)测定.土壤充水孔隙(soil water-filled pore space, WFPS)由下列方程计算获得:
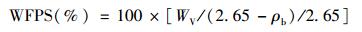
|
(1) |
式中, WV是体积含水量, ρb是土壤容重, 2.65是土壤颗粒密度.
1.5 数据分析通过差减法计算凋落物呼吸:
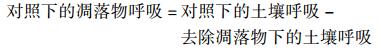
|
(2) |
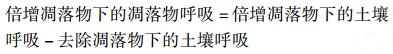
|
(3) |
利用差减法会导致个别观测日的凋落物呼吸出现负值(10%), 可能由于土壤呼吸相比凋落物控制处理而言本身具有较高的变异性、或者地表凋落物对土壤呼吸的拟制作用、甚至是小区自身具有较大的空间异质性等原因所致[9, 31].
利用指数和Q10关系模型拟合土壤温度对凋落物呼吸(去除负的凋落物呼吸)的影响[9]:
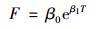
|
(4) |
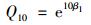
|
(5) |
式中, F[μmol·(m2·s)-1]为凋落物呼吸, T(℃)为5 cm处的土壤温度, β0和β1是拟合参数.
凋落物呼吸对土壤水分的响应机制十分复杂, 这种复杂性表现在以下两个方面[9, 16]:① 一般而言凋落物呼吸在过高或者过低的土壤水分条件下均会受到不同程度的抑制, 因此利用抛物线模型拟合土壤水分对凋落物呼吸的影响; ② 土壤水分对凋落物呼吸的影响很有可能混淆了土壤温度对其的影响.
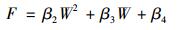
|
(6) |
式中, W是0~5 cm处的土壤水分, β2、β3和β4是拟合系数.
土壤温度和土壤水分的交互作用能很好地拟合二者分对凋落物呼吸的影响, 因此利用双变量模型模拟凋落物呼吸对土壤温度和土壤水分的响应机制[16].
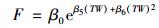
|
(7) |
式中, β5和β6是拟合系数.
利用线性内插法估算累积土壤呼吸/凋落物呼吸, 对于个别观测日出现负的地表凋落物呼吸在计算累积凋落物呼吸时该具体观测日的凋落物呼吸按照0计算[9, 16].利用SigmaPlot 10.0软件的线性回归模型拟合年平均土壤水分和地表凋落物量对凋落物呼吸温度敏感性的影响.
地表凋落物量和年平均土壤水分对凋落物呼吸温度敏感性的交互影响由下列方程获得[9]:
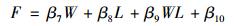
|
(8) |
式中, F为凋落物呼吸温度敏感性, W为年平均土壤水分, L为地表凋落物量, β7、β8、β9和β10是拟合系数.
为了单独量化年平均土壤水分和地表凋落物量对凋落物呼吸温度敏感性的贡献, 在方程(8) 的基础上通过方程(9)、(10) 计算地表凋落物量和年平均土壤水分的标准化回归系数(β), 地表凋落物量和年平均土壤水分β值的大小即可以反映出二者对凋落物呼吸温度敏感性贡献的大小[9]:
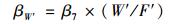
|
(9) |
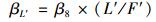
|
(10) |
式中, βW′和βL′分别是年平均土壤水分和地表凋落物量的β值, W′和L′分别是年平均土壤水分和地表凋落物量的标准偏差, F′是拟合的凋落物呼吸温度敏感性的标准偏差.
2 结果与分析 2.1 不同凋落物处理下的土壤微气候差异不同凋落物处理下的土壤温度具有相似的季节变化趋势, 且这种变化趋势与大气温度的变化趋势基本一致, 均呈现出夏季大于其它季节的趋势(图 1).此外, 不同凋落物处理下的平均土壤温度呈现出倍增凋落物处理>对照处理>去除凋落物处理的趋势(14.92℃、14.76℃和14.27℃), 但各处理间的差异没有达到统计显著水平(P>0.05).

|
图 1 不同凋落物处理下的土壤温度和水分 Fig. 1 Soil temperature/moisture for the different surface litter control measures |
不同凋落物处理下的土壤水分具有相似的季节变化趋势, 且这种变化趋势与降雨的不均匀分布密切相关(图 1).此外, 不同凋落物处理下的平均土壤水分差异显著(P<0.05), 且呈现出去除凋落物处理>对照处理>倍增凋落物处理的趋势(41.58% WFPS、38.26% WFPS和35.37% WFPS), 这可能是由于尼龙网的遮阴效果所导致的.
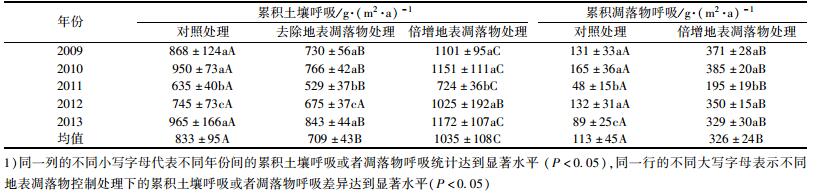
2.2 凋落物呼吸的动态变化趋势不同地表凋落物处理下的土壤呼吸速率具有相似的变化趋势, 且这种变化趋势与大气或者土壤温度的变化趋势基本一致(图 1和2), 均呈现出夏季大于其它季节的趋势(LCK: [4.05 μmol·(m2·s)-1]和[1.91 μmol·(m2·s)-1]; LA: [4.76 μmol·(m2·s)-1]和[2.40 μmol·(m2·s)-1]; LR: [3.50 μmol·(m2·s)-1]和1.76 [μmol·(m2·s)-1]).此外, 不同地表凋落物处理下的土壤呼吸速率呈现出LA>LCK>LR的趋势(表 1:[3.34 μmol·(m2·s)-1]、[2.72 μmol·(m2·s)-1]和[2.41 μmol·(m2·s)-1]; [1 035 g·(m2·a)-1]、[833 g·(m2·a)-1]和[709 g·(m2·a)-1]).

|
图 2 不同地表凋落物处理下的土壤呼吸/凋落物呼吸 Fig. 2 Soil/surface litter respiration for the different surface litter control measures |
凋落物呼吸的动态变化趋势与大气或者土壤温度的变化趋势基本一致(图 1和2), 呈现出夏季大于其他季节的趋势[倍增凋落物处理:1.51 μmol·(m2·s)-1和0.58 μmol·(m2·s)-1; 对照处理:0.79 μmol·(m2·s)-1和0.31 μmol·(m2·s)-1], 且不同地表凋落物处理下的凋落物呼吸速率呈现出倍增凋落物处理>对照处理的趋势[表 1:1.03 μmol·(m2·s)-1和0.51 μmol·(m2·s)-1; 326 g·(m2·a)-1和113 g·(m2·a)-1].
|
|
表 1 不同地表凋落物处理下的累积土壤呼吸/凋落物呼吸 Table 1 Annual cumulative soil/surface litter respirationin for the different surface litter control measures |
2.3 凋落物呼吸的影响因素
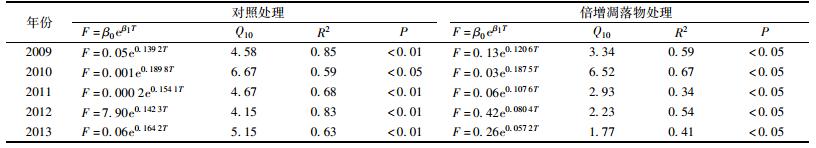
土壤温度对凋落物呼吸的影响用指数关系模型进行拟合, 且土壤温度可以解释59%~85% (对照处理)和34%~67% (倍增凋落物处理)的凋落物呼吸变异性(表 2).
|
|
表 2 土壤温度对凋落物呼吸的影响 Table 2 Effect of soil temperature on surface litter respiration |
土壤水分对凋落物呼吸的影响比较复杂, 这种复杂性表现在凋落物呼吸对土壤水分的响应可能混淆了土壤温度对凋落物呼吸的影响(图 3).在对照处理下, 当土壤温度大于17℃时, 凋落物呼吸对土壤水分的响应可以用抛物线关系模型进行拟合(P<0.05), 且土壤水分可以解释41%的凋落物呼吸变异性, 当土壤温度小于17℃时, 凋落物呼吸不受土壤水分的制约(P>0.05);在倍增凋落物处理下, 无论是土壤温度大于17℃或者小于17℃, 凋落物呼吸均不同程度受到土壤水分的影响(P<0.05), 在相对高温时土壤水分可以解释高达42%的凋落物呼吸变异性(P<0.05), 在相对低温时土壤水分仅可以解释18%的凋落物呼吸变异性(P<0.05).除了土壤温度和土壤水分以外, 凋落物呼吸对土壤温度和土壤水分的响应曲面可以直观明了地反映出土壤温度和土壤水分以及二者交互作用对凋落物呼吸的影响(图 4, P<0.01).

|
图 3 土壤水分对凋落物呼吸的影响 Fig. 3 Effect of soil moisture on surface litter respiration |

|
图 4 土壤水分和土壤温度的交互作用对凋落物呼吸的影响 Fig. 4 Effect of the interplay between soil temperature and soil moisture on surface litter respiration |
在季节尺度上(一个完整的生长季), 土壤温度是影响凋落物呼吸的主要因素, 且凋落物呼吸温度敏感性的年际差异显著(P<0.05):在对照处理下, 凋落物呼吸温度敏感性的最小值为4.15 (2012年), 最大值为6.67 (2010年), 均值为5.10, 变异系数为19% (表 2); 在倍增凋落物处理下, 其变化于1.17~6.52之间, 均值为3.36, 变异系数为56% (表 2).此外, 倍增地表凋落物显著地减少了凋落物呼吸温度敏感性(P<0.05), 平均倍增地表凋落物处理减少了34%的凋落物呼吸温度敏感性(3.36和5.10).
在年际尺度上, 凋落物呼吸温度敏感性与年平均土壤水分、地表凋落物量以及二者的交互作用密切相关(表 3, P<0.01).同时进一步的研究发现(图 5), 对凋落物呼吸温度敏感性的贡献(B值)呈现出土壤水分大于地表凋落物量的趋势(对照处理: 2.68和2.04;倍增凋落物处理:1.37和0.69).此外, 进一步的研究结果发现:地表凋落物量主要取决于夏季降雨(图 6), 且对数关系可以很好地模拟夏季降雨对地表凋落物量的影响(y=303lnx-1 228, R2=0.96, P<0.01).
|
|
表 3 地表凋落物量(L)和年平均土壤水分(W)的交互作用对地表凋落物呼吸温度敏感性的影响 Table 3 Effect of the interplay between surface litter biomass (L) and annual mean soil moisture (W) on the temperature sensitivity of surface litter respiration |


|
图 5 地表凋落物量和土壤水分对凋落物呼吸温度敏感性的贡献 Fig. 5 Contribution of surface litter biomass and soil moisture on the temperature sensitivity of surface litter respiration |

|
图 6 夏季降雨对凋落物量的影响 Fig. 6 Effect of summer rainfall on surface litter biomass |
在本研究中, 不同地表凋落物控制处理下的累积凋落物呼吸变化于48~385 g·(m2·a)-1之间, 其正好位于全球林地累积地表凋落物呼吸的变化范围[61~970 g·(m2·a)-1]之内[2, 7, 10], 然而本研究中的平均累积凋落物呼吸却低于其它拥有充足降雨的林地生态系统的平均累积凋落物呼吸[8, 27].此外, 不同地表凋落物控制处理下的凋落物呼吸温度敏感性变化于1.17~6.67之间, 其恰好也位于全球林地地表凋落物呼吸温度敏感性的变化范围(1.29~10.6) 之内[7, 16, 18, 19].但是, 本研究中的平均凋落物呼吸温度敏感性却高于其它林地生态系统的平均凋落物呼吸温度敏感性(4.23和3.61)[7, 16, 18, 19], 这可能是因为黄土高原地区是我国典型的生态脆弱区以及典型的干旱和半干旱地区, 其有限的降雨(544 mm)和较低的地表凋落物归还量[477 g·(m2·a)-1]均限制了该区的地下生理过程以及造成该区的生态系统对气候变化相对较为敏感[9, 16].
不同地表凋落物控制处理下的凋落物呼吸温度敏感性的年际变化差异显著(P<0.05), 而土壤水分和地表凋落物是驱动其变化的主要因素, 究其原因可能与以下因素有关:① 在水分匮乏的干旱、半干旱的黄土高原地区, 土壤水分是控制地下生理过程的主要限制性因素[32, 33], 因此土壤水分的微小波动必然会影响地下生理过程的相应变化[34, 35].例如, 中国科学院海北高寒草甸生态系统定位研究站的室内培养试验研究结果表明, 青藏高原高寒矮嵩草草甸生态系统的凋落物呼吸温度敏感性随着土壤湿度的增加呈现出增加的趋势[20].在本研究中, 凋落物呼吸温度敏感性随着土壤水分的增加呈现出线性增加的趋势(y=0.08x+1.74, R2=0.42, P<0.05), 这与已有的研究结果相一致[20]; ② 降雨是黄土高原地区唯一的水分来源, 降雨的剧烈波动不可避免地影响到地表凋落物量[21, 22].在本研究中, 地表凋落物量随着夏季降雨的增加呈现出对数增加的趋势(图 6), 这与已有的研究结果相类似[36].除了土壤水分以外, 底物的数量(地表凋落物量)也是影响凋落物呼吸温度敏感性的重要因素[18].例如, 在湖北省赤壁市丁母山林场的纯毛竹林中的研究发现, 凋落物呼吸温度敏感性随着地表凋落物归还量的减少(因复垦和劈草造成归还到土壤中的地表凋落物量不同程度地降低)而呈现出降低趋势[18].且研究结果显示, 凋落物呼吸温度敏感性随着地表凋落物量的增加而呈线性增加趋势(y=0.01x+0.05, R2=0.58, P<0.05), 这与已有的研究结果相匹配[18].
3.2 应用意义地表凋落物控制措施一方面通过直接影响微生物进行新陈代谢的食物来源(地表凋落物), 另一方面通过间接影响土壤微环境差异影响地表凋落物呼吸温度敏感性[18, 20].本研究结果表明, 当地表凋落物量倍增以后, 累积凋落物呼吸量增加了将近3倍, 但是相应地凋落物呼吸温度敏感性却降低了34% (3.36和5.10), 即在未来持续增温的条件下, 倍增地表凋落物条件下的凋落物呼吸对升温相对不敏感.究其原因可能与以下因素有关:① 倍增地表凋落物量以后由于地表凋落物层的加厚很有可能拟制了土壤中CO2的排放[27]; ② 倍增地表凋落物后很有可能通过增加土壤水分或者减少土壤温度降低凋落物呼吸温度敏感性[37], 这是因为已有的研究结果表明, 地表凋落物呼吸温度敏感性随着土壤温度的增加或者土壤水分的减少而呈现出减少的趋势[18, 20]; ③ 刺槐林属于固氮树种, 与其它非固氮树种相比较而言, 刺槐林地表凋落物具有相对较低的C:N[38], 而研究结果显示相对活性的底物具有较小的呼吸温度敏感性[39, 40], 但具体的原因有待进一步的试验.
此外, 在对照处理下, 地表凋落物碳输入量远大于其输出量[图 7:215 g·(m2·a)-1和113 g·(m2·a)-1], 即大约有超过50%的地表凋落物碳滞留在该刺槐林生态系统中.但在倍增处理下, 仅有24%的地表凋落物碳滞留在该刺槐林生态系统中[图 7:430 g·(m2·a)-1和326 g·(m2·a)-1], 即在该人工刺槐林生态系统中地表凋落物的增加未必意味着土壤有机碳储量的增加.

|
图 7 不同凋落物处理下的土壤碳平衡状况 Fig. 7 Balance of soil carbon for the different surface litter control measures |
在黄土高原地区的人工刺槐林生态系统中, 以凋落物呼吸的形式释放到大气中CO2大约为113 g·(m2·a)-1, 约占总土壤呼吸的15%.本研究结果显示, 凋落物呼吸温度敏感性的年际变化主要取决于年平均土壤水分和地表凋落物量, 而年平均土壤水分和地表凋落物量均受降雨频率与分布的制约(图 6和8).因此, 在旱地生态系统中, 为了更加准确地预测凋落物呼吸的动态变化趋势, 降雨的剧烈波动性是必须考虑的一个重要因素之一.

|
图 8 降雨频率与分布对土壤水分的影响 Fig. 8 Effect of rainfall frequency and distribution on annual mean soil moisture |
研究凋落物呼吸温度敏感性的变化特征及其影响因素对丰富和准确地理解地区的土壤碳循环具有重要意义.凋落物呼吸温度敏感性的年际差异显著(P<0.05), 而这种年际差异性主要取决于年平均土壤水分、地表凋落物量以及二者的交互作用(P<0.01), 且对凋落物呼吸温度敏感性的贡献呈现出土壤水分大于地表凋落物量的趋势.此外, 在倍增凋落物条件下, 凋落物呼吸增加了将近3倍, 但是相应的凋落物呼吸温度敏感性却减少了34%.无论是在对照或者是在倍增凋落物处理下, 均呈现出地表凋落物的输入量大于输出量的趋势.因此, 凋落物控制措施、土壤水分、地表凋落物量是黄土区刺槐林凋落物呼吸温度敏感性的主要驱动因素.
致谢: 感谢西北农林科技大学水土保持研究所郭胜利课题组、中国科学院长武黄土高原农业生态试验站和宝鸡文理学院张蓓蓓副教授给予的帮助.| [1] | Kuzyakov Y. Sources of CO2 efflux from soil and review of partitioning methods[J]. Soil Biology and Biochemistry, 2006, 38(3): 425-448. DOI:10.1016/j.soilbio.2005.08.020 |
| [2] | Raich J W, Schlesinger W H. The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate[J]. Tellus B: Chemical and Physical Meteorology, 1992, 44(2): 81-99. DOI:10.3402/tellusb.v44i2.15428 |
| [3] | Prescott C E. Litter decomposition: what controls it and how can we alter it to sequester more carbon in forest soils?[J]. Biogeochemistry, 2010, 101(1-3): 133-149. DOI:10.1007/s10533-010-9439-0 |
| [4] | Xu S, Liu L, Sayer E J. Variability of aboveground litter inputs alters soil physicochemical and biological processes: a meta-analysis of litterfall-manipulation experiments[J]. Biogeosciences Discussions, 2013, 10(3): 5245-5272. DOI:10.5194/bgd-10-5245-2013 |
| [5] | Kim H, Hirano T, Koike T, et al. Contribution of litter CO2 production to total soil respiration in two different deciduous forests[J]. Phyton-Annales Rei Botanicae, 2005, 45(4): 385-388. |
| [6] | Liang N, Hirano T, Zheng Z M, et al. Soil CO2 efflux of a larch forest in northern Japan[J]. Biogeosciences, 2010, 7(11): 3447-3457. DOI:10.5194/bg-7-3447-2010 |
| [7] | Rey A, Pegoraro E, Tedeschi V, et al. Annual variation in soil respiration and its components in a coppice oak forest in Central Italy[J]. Global Change Biology, 2002, 8(9): 851-866. DOI:10.1046/j.1365-2486.2002.00521.x |
| [8] | Sulzman E W, Brant J B, Bowden R D, et al. Contribution of aboveground litter, belowground litter, and rhizosphere respiration to total soil CO2 efflux in an old growth coniferous forest[J]. Biogeochemistry, 2005, 73(1): 231-256. DOI:10.1007/s10533-004-7314-6 |
| [9] | Zhang Y J, Gan Z T, Li R J, et al. Litter production rates and soil moisture influences interannual variability in litter respiration in the semi-arid Loess Plateau, China[J]. Journal of Arid Environments, 2016, 125: 43-51. DOI:10.1016/j.jaridenv.2015.09.016 |
| [10] | Wang Y D, Wang H M, Ma Z Q, et al. Contribution of aboveground litter decomposition to soil respiration in a subtropical coniferous plantation in Southern China[J]. Asia-Pacific Journal of Atmospheric Sciences, 2009, 45(2): 137-147. |
| [11] | Zimmermann M, Meir P, Bird M, et al. Litter contribution to diurnal and annual soil respiration in a tropical montane cloud forest[J]. Soil Biology and Biochemistry, 2009, 41(6): 1338-1340. DOI:10.1016/j.soilbio.2009.02.023 |
| [12] |
邓琦, 刘世忠, 刘菊秀, 等. 南亚热带森林凋落物对土壤呼吸的贡献及其影响因素[J]. 地球科学进展, 2007, 22(9): 976-986. Deng Q, Liu S Z, Liu J X, et al. Contributions of litter-fall to soil respiration and its affacting factors in southern subtropical forests of China[J]. Advances in Earth Science, 2007, 22(9): 976-986. |
| [13] |
赵昕, 张万军, 沈会涛, 等. 针阔树种人工林地表凋落物对土壤呼吸的贡献[J]. 中国生态农业学报, 2014, 22(11): 1318-1325. Zhao X, Zhang W J, Shen H T, et al. Contributions of aboveground litter to soil respiration in coniferous and deciduous plantations[J]. Chinese Journal of Eco-Agriculture, 2014, 22(11): 1318-1325. |
| [14] |
周小刚, 郭胜利, 车升国, 等. 黄土高原刺槐人工林地表凋落物对土壤呼吸的贡献[J]. 生态学报, 2012, 32(7): 2150-2157. Zhou X G, Guo S L, Che S G, et al. Aboveground litter contribution to soil respiration in a black locust plantation in the Loess Plateau[J]. Acta Ecologica Sinica, 2012, 32(7): 2150-2157. |
| [15] |
王丽丽, 宋长春, 郭跃东, 等. 三江平原不同土地利用方式下凋落物对土壤呼吸的贡献[J]. 环境科学, 2009, 30(11): 3130-3135. Wang L L, Song C C, Guo Y D, et al. Contribution of litter to soil respiration under different land-use types in Sanjiang plain[J]. Environmental Scinece, 2009, 30(11): 3130-3135. DOI:10.3321/j.issn:0250-3301.2009.11.002 |
| [16] | Zhang Y J, Guo S L, Liu Q F, et al. Influence of soil moisture on litter respiration in the semiarid Loess Plateau[J]. PLoS One, 2014, 9(12): e114558. DOI:10.1371/journal.pone.0114558 |
| [17] |
田祥宇, 涂利华, 胡庭兴, 等. 华西雨屏区苦竹人工林土壤呼吸各组分特征及其温度敏感性[J]. 应用生态学报, 2012, 23(2): 293-300. Tian X Y, Tu L H, Hu T X, et al. Characteristics of soil respiration components and their temperature sensitivity in a Pleioblastus amarus plantation in rainy area of west China[J]. Chinese Journal of Applied Ecology, 2012, 23(2): 293-300. |
| [18] |
范少辉, 唐晓鹿, 漆良华, 等. 不同经营措施对毛竹林土壤呼吸温度敏感性的影响[J]. 四川农业大学学报, 2012, 30(3): 300-307. Fan S H, Tang X L, Qi L H, et al. Effects of different managements on temperature sensitivity of soil respiration of Phyllostachy edulis forest[J]. Journal of Sichuan Agricultural University, 2012, 30(3): 300-307. |
| [19] |
武传胜, 沙丽清, 张一平. 哀牢山中山湿性常绿阔叶林凋落物对土壤呼吸及其温度敏感性的影响[J]. 东北林业大学学报, 2012, 40(6): 37-40. Wu C S, Sha L Q, Zhang Y P. Effect of litter on soil respiration and its temperature sensitivity in a montane evergreen broad-leaved forest in Ailao mountains, Yunnan[J]. Journal of Northeast Forestry University, 2012, 40(6): 37-40. |
| [20] |
苏爱玲. 气候变化对高寒草甸枯落物分解特性的影响及温度敏感性研究[D]. 西宁: 中国科学院西北高原生物研究所, 2010. Su A L. Effect of climate change on litter decomposition and temperature sensitivity in Alpine Meadow on Qinghai-Tibetan Plateau[D]. Xi'ning: Northwest Institute of Plateau Biology, Chinese Academy of Sciences, 2010. http://ir.nwipb.ac.cn/handle/363003/3418?mode=full&submit_simple=Show+full+item+record |
| [21] | Austin A T. Differential effects of precipitation on production and decomposition along a rainfall gradient in Hawaii[J]. Ecology, 2002, 83(2): 328-338. |
| [22] | Travers S K, Eldridge D J. Increased rainfall frequency triggers an increase in litter fall rates of reproductive structures in an arid eucalypt woodland[J]. Austral Ecology, 2013, 38(7): 820-830. DOI:10.1111/aec.2013.38.issue-7 |
| [23] | Boone R D, Nadelhoffer K J, Canary J D, et al. Roots exert a strong influence on the temperature sensitivity of soil respiration[J]. Nature, 1998, 396(6711): 570-572. DOI:10.1038/25119 |
| [24] | Karhu K, Fritze H, Hämäläinen K, et al. Temperature sensitivity of soil carbon fractions in boreal forest soil[J]. Ecology, 2010, 91(2): 370-376. DOI:10.1890/09-0478.1 |
| [25] | Zhang Y J, Guo S L, Liu Q F, et al. Responses of soil respiration to land use conversions in degraded ecosystem of the semi-arid Loess Plateau[J]. Ecological Engineering, 2015, 74: 196-205. DOI:10.1016/j.ecoleng.2014.10.003 |
| [26] | Shi W Y, Zhang J G, Yan M J, et al. Seasonal and diurnal dynamics of soil respiration fluxes in two typical forests on the semiarid Loess Plateau of China: temperature sensitivities of autotrophs and heterotrophs and analyses of integrated driving factors[J]. Soil Biology and Biochemistry, 2012, 52: 99-107. DOI:10.1016/j.soilbio.2012.04.020 |
| [27] | Yan W D, Chen X Y, Tian D L, et al. Impacts of changed litter inputs on soil CO2 efflux in three forest types in central south China[J]. Chinese Science Bulletin, 2013, 58(7): 750-757. DOI:10.1007/s11434-012-5474-7 |
| [28] | Valentini C M A, Sanches L, de Paula S R, et al. Soil respiration and aboveground litter dynamics of a tropical transitional forest in northwest Mato Grosso, Brazil[J]. Journal of Geophysical Research-Biogeosciences, 2008, 113(G1): G00B10. |
| [29] |
高会议, 郭胜利, 刘文兆. 黄土旱塬裸地土壤呼吸特征及其影响因子[J]. 生态学报, 2011, 31(18): 5217-5224. Gao H Y, Guo S L, Liu W Z. Characteristics of soil respiration in fallow and its influencing factors at arid-highland of Loess Plateau[J]. Acta Ecologica Sinica, 2011, 31(18): 5217-5224. |
| [30] |
周洪华, 李卫红, 杨余辉, 等. 干旱区不同土地利用方式下土壤呼吸日变化差异及影响因素[J]. 地理科学, 2011, 31(2): 190-196. Zhou H H, Li W H, Yang Y H, et al. Soil respiration variant and its effecting factors at different land use in Arid land[J]. Scientia Geographica Sinica, 2011, 31(2): 190-196. |
| [31] | Berryman E M, Marshall J D, Kavanagh K. Decoupling litter respiration from whole-soil respiration along an elevation gradient in a Rocky Mountain mixed-conifer forest[J]. Canadian Journal of Forest Research, 2014, 44(5): 432-440. DOI:10.1139/cjfr-2013-0334 |
| [32] | Reth S, Reichstein M, Falge E. The effect of soil water content, soil temperature, soil pH-value and the root mass on soil CO2 efflux-a modified model[J]. Plant and Soil, 2005, 268(1): 21-33. DOI:10.1007/s11104-005-0175-5 |
| [33] | Wang B, Zha T S, Jia X, et al. Soil moisture modifies the response of soil respiration to temperature in a desert shrub ecosystem[J]. Biogeosciences, 2014, 11(2): 259-268. DOI:10.5194/bg-11-259-2014 |
| [34] | Bowden R D, Newkirk K M, Rullo G M. Carbon dioxide and methane fluxes by a forest soil under laboratory-controlled moisture and temperature conditions[J]. Soil Biology and Biochemistry, 1998, 30(12): 1591-1597. DOI:10.1016/S0038-0717(97)00228-9 |
| [35] | Zhang Y J, Guo S L, Zhao M, et al. Soil moisture influence on the interannual variation in temperature sensitivity of soil organic carbon mineralization in the Loess Plateau[J]. Biogeosciences, 2015, 12(11): 3655-3664. DOI:10.5194/bg-12-3655-2015 |
| [36] | Lawrence D, Foster D. Changes in forest biomass, litter dynamics and soils following shifting cultivation in southern Mexico: an overview[J]. Interciencia, 2002, 27(8): 400-408. |
| [37] | Fekete I, Varga C, Biró B, et al. The effects of litter production and litter depth on soil microclimate in a central european deciduous forest[J]. Plant and Soil, 2016, 398(1-2): 291-300. DOI:10.1007/s11104-015-2664-5 |
| [38] |
郭胜利, 马玉红, 车升国, 等. 黄土区人工与天然植被对凋落物量和土壤有机碳变化的影响[J]. 林业科学, 2009, 45(10): 14-18. Guo S L, Ma Y H, Che S G, et al. Effects of artificial and natural vegetations on litter production and soil organic carbon change in loess hilly areas[J]. Scientia Silvae Sinicae, 2009, 45(10): 14-18. DOI:10.11707/j.1001-7488.20091003 |
| [39] | Conant R T, Drijber R A, Haddix M L, et al. Sensitivity of organic matter decomposition to warming varies with its quality[J]. Global Change Biology, 2008, 14(4): 868-877. DOI:10.1111/gcb.2008.14.issue-4 |
| [40] | Wetterstedt J Å M, Persson T, Ågren G I. Temperature sensitivity and substrate quality in soil organic matter decomposition: results of an incubation study with three substrates[J]. Global Change Biology, 2010, 16(6): 1806-1819. |