 2017, Vol. 38
2017, Vol. 38



2. 南京师范大学环境学院, 南京 210023;
3. 江西省红壤研究所, 南昌 331717
2. School of Environmental Sciences, Nanjing Normal University, Nanjing 210023, China;
3. Jiangxi Institute of Red Soil, Nanchang 331717, China
土壤氮素地球化学循环是农业生态系统研究的重要内容, 硝化作用是土壤氮素转化的重要过程, 是全球氮素循环中心, 分为自养硝化和异养硝化[1].自养硝化是指自养硝化微生物以无机碳为碳源将NH3氧化为亚硝酸盐或者硝酸盐的过程, 异养硝化是指异养硝化微生物以有机碳或有机氮化合物为碳源和能源进行硝化作用的过程.我国典型生态系统中, 农田硝化活性较高[2], 且以自养硝化作用为主[3].自养硝化包括氨氧化和亚硝酸盐氧化过程, 其中氨氧化过程是限速步骤, 由氨氧化微生物执行, 包括氨氧化古菌(ammonia-oxidizing archaea, AOA)和细菌(ammonia-oxidizing bacteria, AOB).氨氧化微生物的生长和活性受pH[4]、底物浓度[5]、温度[6]、土壤类型[7]、氧气含量[8]等多种环境因子影响.施肥是农田生态系统重要管理措施, 化肥和有机肥料输入可提升土壤营养元素含量、改变土壤C/N, 施有机肥可增加土壤pH值、提高土壤通透性等, 而施化肥可引起土壤酸化、板结化等问题[9].施肥后土壤性质改变会影响土壤微生物群落及生态功能[10].目前, 对于中性和偏酸性土壤中施肥对氨氧化微生物影响的研究较多, 发现施加化肥和有机肥可提升土壤AOA和AOB丰度, 其中主要AOA类群为Nitrosophara, 主要AOB类群为Nitrosospira[11, 12]; 且在中性和碱性土壤中AOB活性占据优势[13, 14].
酸碱度处于4.5~5.5之间的铁铝酸性红壤约占全国土地总面积22.7%, 具有氧化势高、酸性强、营养贫瘠等特点.施肥对于提高酸性红壤农业产量及经济效益尤为重要.由于红壤本底pH较低, 施化肥对土壤造成的酸化作用不再明显, 而施有机肥则显著提升土壤pH、养分含量及通气性[15].研究发现在强酸性土壤(pH<5.5) 中, 对低底物浓度有更强亲和力的AOA活性更高[16, 17].但目前酸性土壤氨氧化微生物对于施肥的响应研究结论尚不一致[15, 18].因此, 施化肥或有机肥对酸性土壤中氨氧化微生物的影响可能与中性或碱性土壤不同, 其环境因子驱动机制也不清楚.
长期定位施肥试验是研究氨氧化微生物生态位分异特征的理想体系.本研究选择长期定位施肥酸性旱地土壤, 构建添加尿素(长期施肥试验用氮肥)短期培养试验, 采用微域培养联用定量PCR(quantitative PCR, qPCR)、变性梯度凝胶电泳(denaturing gradient gel electrophoresis, DGGE)及Illumina MiSeq高通量测序等分子生态学技术, 分析培养过程中发挥硝化活性的、且占据活性优势的硝化微生物类群, 研究自养硝化微生物群落及活性特征, 并认知其与土壤理化因子间的关系, 明确长期施用化肥或有机肥对酸性旱地土壤硝化活性及自养硝化微生物的影响及其机制, 以期为认识自养硝化微生物生理代谢多样性及生态功能提供了试验数据, 也为酸性旱地土壤施肥管理提供参考.
1 材料与方法 1.1 取样地点及土样处理供试土壤采集于江西进贤红壤生态试验站(28°21′04″N, 116°10′16″E), 该地区气候为典型亚热带季风气候, 全年平均气温为17.7℃, 年降水量1 727 mm.试验地建于1986年, 设有种植双季玉米的长期定位施肥试验田, 种植期为每年4~11月.本研究选用不施肥(CK)、施化肥(NPK)和施有机肥(OM)这3种土壤, 各设置3个重复小区(5.5 m×4 m).肥料年施用量:氮肥(尿素), 120 kg·hm-2; 磷肥(过磷酸钙), 60 kg·hm-2; 钾肥(氯化钾), 120 kg·hm-2; 有机肥(熟猪粪), 2 000 kg·hm-2.有机肥和无机肥于每年玉米种植前作为基肥一次性施加.于2015年4月(早季玉米种植前)采样, 各小区按S型五点采样法, 采集耕作表层(0~15 cm)土壤, 去除杂物与根系过2 mm筛后混匀.部分土壤立即储存于-80℃用于分子生物学分析, 部分土壤风干后用于理化性质分析, 其余土壤于4℃保存.本研究土壤基本理化性质测定方法参照文献[19], 其中pH分别采用去离子水和2 mol·L-1 KCl溶液以水土比2.5:1浸提.
1.2 土壤微域培养及样品采集称取10.00 g鲜土于120 mL血清瓶中, 橡胶塞封口, 于28℃避光预培养2 d.预培养结束后加入终浓度(以干土计)为100 mg·kg-1的Urea(尿素)溶液并添加去离子水调节土壤至60%最大持水量, 用于筛选土壤中发挥硝化活性的功能微生物类群.微域培养设置添加和未添加C2H2(1%)处理, 用于区分自养和异养硝化过程[20].每种处理设置12个血清瓶重复.密封后于28℃避光培养7 d.分别于0、1、3、7 d破坏性采样, 每次3个血清瓶重复.
1.3 土壤总DNA提取及amoA基因丰度测定土壤微生物总DNA采用FastDNA® SPIN Kit for Soil(MP, Biomedicals, USA)试剂盒提取. amoA基因丰度采用qPCR法在CFX96 Real-Time System(BIO-RAD, Laboratories Inc, USA)上测定. AOA、AOB的amoA基因扩增引物分别为Arch-amoAF (5′-STA ATG GTC TGG CTT AGA CG-3′)/Arch-amoAR (5′-GCG GCC ATC CAT CTG TAT GT-3′)[21]和amoA-1F (5′-GGG GTT TCT ACT GGT GGT-3′)/amoA-2R (5′-CCC CTC KGS AAA GCC TTC TTC-3′)[22]. qPCR反应体系、扩增程序、熔解曲线程序及PCR抑制检测方法见文献[15].采用10倍稀释含有标靶基因的重组质粒制作标准曲线, AOA、AOB的amoA基因标准曲线模板浓度分别为2.44×109~2.44×103 copies·μL-1和1.93×109~1.93×103 copies·μL-1, 扩增效率分别为82.4%(R2=0.998) 和91.8%(R2=0.997).
1.4 氨氧化微生物的PCR-DGGE分析AOA amoA基因扩增引物为Arch-amoAF/Arch-amoAR. PCR扩增采用TaKaRa Ex Taq HS(Takara, Japan)热启动酶, 反应体系同上.扩增程序为:预变性95℃ 3 min, 变性95℃ 30 s, 退火55℃ 30 s, 延伸72℃ 30 s, 设置35个循环, 终延伸72℃ 6 min.对PCR产物进行1.2%琼脂糖凝胶电泳检测及凝胶成像(ChemiDOC XRS+, BIO-RAD, USA). AOB扩增产物浓度过低(<0.5 ng·μL-1), 不适用于后续DGGE分析.
每个样品取约200 ng PCR产物在Dcode系统(Dcode Universal Mutation Detection System, BIO-RAD, USA)中进行DGGE分析.聚丙烯酰胺凝胶浓度为6%, 变性梯度为30%~65%(丙烯酰胺:聚丙烯酰胺=37.5:1, 100%变性剂由40%去离子甲酰胺和7 mol·L-1的尿素配制而成). 60℃, 80 V, 电泳17 h.采用SYBR Green I(Invitrogen, USA)染色30 min后成像.切割下DGGE图谱中优势条带, 清洗后加入40 μL无菌水于4℃静置过夜以洗脱DNA.采用pEASY-T1 vector(TransGen Biotech, China)进行克隆, 每个条带选取3个阳性克隆子送测(Sangon Biotech, China).将测序结果与NCBI数据库进行比对.
1.5 MiSeq高通量测序分析16S rRNA基因扩增引物采用V4-V5高变区通用引物515F (5′-GTG CCA GCM GCC GCG GTA A-3′)/907R (5′-CCG TCA ATT CCT TTR AGT TT-3′).反应体系为50 μL, 包括25 μL TaKaRa Ex Taq HS (Takara, Japan), 20 μmol·L-1上下游引物各0.5 μL, 1 μL模板DNA及23 μL无菌水.反应程序:预变性94℃ 3 min, 变性95℃ 30 s, 退火55℃ 30 s, 延伸72℃ 30 s, 设置30个循环, 终延伸72℃ 6 min.设置3个重复, 对PCR产物进行1.5%琼脂糖凝胶电泳检测及割胶纯化(Agarose Gel DNA Extraction Kit, Takara, Japan).委托中科院南京土壤研究所分析测试中心采用Illumina MiSeq平台进行高通量测序分析.
采用FLASH(v1.2.7, http://ccb.jhu.edu/software/FLASH/)对序列进行拼接; 通过Qiime(V1.8.0, http://qiime.org/)和USEARCH(V5.2.236, http://www.drive5.com/usearch/)过滤序列并去除嵌合体.共获得228 031条序列, CK、NPK和OM土壤获得的高质量序列数分别为26 040~27 730、25 482~26 907和21 687~25 220条.为避免测序深度不同导致的误差, 每个样本随机抽取19 600条序列参与统计分析.以97%的序列相似度水平划分OTU, 剔除相对丰度低于0.001%的OTU.序列与RDP数据库(http://rdp.cme.msu.edu/)进行比对分类.
1.6 数据分析本研究以添加自养硝化作用抑制剂C2H2处理的硝化速率表征异养硝化速率, 以不添加C2H2与添加C2H2处理的硝化速率差值表征自养硝化速率.采用Excel 2013、SPSS 16.0统计软件对试验数据进行分析; 采用单因素方差分析法区分样品间的显著性差异(n=3, Duncan'test, P<0.05);采用MEGA 4.0构建系统发育树, 建树方法为Neighbor-Joining法, Bootstrap设为1 000.采用逐步回归分析方法分析土壤硝化活性、硝化微生物丰度及组成与土壤理化因子之间的关系, 其中pH值选用KCl溶液浸提测定数值.
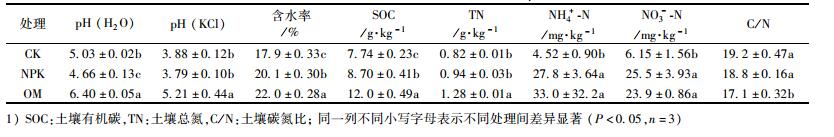
2 结果与分析 2.1 长期施肥处理土壤基本理化性质单因素方差分析表明(表 1), 与CK土壤相比, NPK土壤含水率、SOC、NH4+-N和NO3--N均显著升高, OM土壤pH、含水率、SOC、TN、NH4+-N和NO3--N均显著升高, C/N显著降低.其中, 用去离子水浸提测得pH值比用2 mol·L-1KCl溶液浸提测定结果高0.87~1.19单位.结果显示施用化肥和有机肥显著提高了土壤含水率、SOC和土壤无机氮含量; 施用有机肥显著提高了土壤pH和TN含量, 但显著降低了土壤C/N.
|
|
表 1 供试土壤理化性质 Table 1 Characteristics of the soil in this study |
2.2 微域培养过程中土壤无机氮含量变化
微域培养过程中, 各处理土壤NH4+-N和NO3--N含量动态变化曲线见图 1.不添加C2H2处理的CK、NPK和OM土壤NO3--N含量显著上升, 分别从第0 d的4.82、24.04、29.14 mg·kg-1上升至第7 d的20.91、67.14、133.15 mg·kg-1; 添加C2H2后, CK土壤NO3--N含量无显著变化, NPK和OM土壤NO3--N含量略微上升, 分别从第0 d的24.15 mg·kg-1和29.12 mg·kg-1上升至第7 d的36.53 mg·kg-1和52.81 mg·kg-1; 土壤NH4+-N含量下降趋势与土壤NO3--N含量上升趋势吻合, 表明土壤NO3--N产生自NH4+-N的氧化, 且7 d培养周期内底物充足.

|
图 1 培养过程中土壤无机氮含量变化规律 Fig. 1 Changes in inorganic nitrogen contents during the incubation |
CK、NPK和OM土壤的自养硝化速率分别为1.86、4.46和11.98 mg·(kg·d)-1, 且OM、NPK土壤自养硝化速率分别是CK土壤的6.44倍和2.40倍; 异养硝化速率分别为0.32、1.60和2.93 mg·(kg·d)-1; 自养硝化占总硝化速率的比例分别为85.32%、73.60%和80.35%(图 2).表明本研究土壤中自养硝化作用占据主导, 施化肥和有机肥显著提高土壤自养硝化速率, 施有机肥提高土壤异养硝化速率.

|
图 2 土壤自养和异养硝化速率 Fig. 2 Soil autotrophic and heterotrophic nitrification rates |
培养前后CK、NPK和OM土壤古菌和细菌的amoA基因拷贝数见图 3(a)和3(b).无论是否添加C2H2处理, CK与NPK土壤培养结束时的AOA和AOB的amoA基因拷贝数与培养前无显著差异; OM土壤不添加C2H2处理(Day-7) 的AOA和AOB的amoA基因拷贝数显著高于培养前(Day-0).即培养过程中CK、NPK的AOA和AOB的绝对丰度并无显著变化, 而OM中AOA和AOB绝对丰度显著增长.培养后施有机肥土壤AOA和AOB的amoA基因绝对丰度(以干土计, 下同)分别为2.04×109 copies·g-1、4.31×108 copies·g-1, 远高于施化肥土壤的5.96×108 copies·g-1、3.75×107 copies·g-1和不施肥土壤的6.58×108 copies·g-1、1.13×107 copies·g-1.

|
图 3 培养前后土壤AOA和AOB amoA基因绝对丰度及16S rRNA基因相对丰度变化规律 Fig. 3 Abundance of archaeal and bacterial amoA genes and relative abundance of AOA-like 16S rRNA and AOB-like 16S rRNA genes before and after the incubation |
培养前后CK、NPK和OM土壤AOA-like 16S rRNA和AOB-like 16S rRNA基因相对丰度(Illumina MiSeq分析)见图 3(c)和3(d).微域培养后(Day-7), 不添加C2H2处理的CK、NPK和OM土壤中AOA-like 16S rRNA的相对丰度分别为1.42%、1.95%和1.87%, 显著高于培养前和添加C2H2处理.不添加C2H2处理, 培养后CK和NPK土壤中AOB-like 16S rRNA的相对丰度分别为1.22%和1.12%, 与培养前和添加C2H2处理无显著差异; 而培养后OM土壤AOB-like 16S rRNA的相对丰度比例为2.20%, 显著高于培养前和添加C2H2处理.
微域培养前后AOA和AOB的绝对丰度和相对丰度变化规律表明:3种土壤中AOA均有明显活性, 而仅在OM土壤中检测到AOB活性.
2.4 微域培养过程中土壤硝化微生物群落组成变化培养前后CK、NPK和OM土壤的AOA群落结构DGGE图谱见图 4. CK和NPK土壤DGGE图谱较相似, 与OM土壤有明显差异. 7 d培养结束后, CK土壤不添加C2H2处理的A1、A3条带亮度加深, A6条带亮度变浅, C2H2处理的A1、A2、A3条带亮度变浅, A6条带亮度变深; NPK处理不添加C2H2处理的A1、A3条带亮度加深, A6条带亮度变浅, C2H2处理的A1、A3条带亮度变浅, A6条带亮度变深; OM土壤不添加C2H2处理的A4条带亮度变深, A9条带亮度变浅, C2H2处理的A4条带亮度变浅.结果显示培养过程不同AOA类群生长状况不同.对各处理土壤DGGE图谱中的优势条带进行测序并进行古菌amoA基因序列系统发育分析(图 5). CK、NPK和OM土壤的DGGE条带A1~A9与已知氨氧化古菌Nitrososphaera sp. EN76和JG1聚成一簇, 均隶属于Nitrososphaera(group 1.1b).培养前后各处理土壤硝化微生物在属水平上的丰度变化见图 6. CK、NPK和OM土壤AOA中的Nitrososphaera相对丰度较培养前均有明显上升, 分别从0.79%、1.28%和1.05%上升至1.41%、1.94%和1.87%, 分别占总AOA类群的99.30%、99.49%和99.99%. Nitrosotalea(Group 1.1a-associated)培养前后相对丰度无显著变化.

|
图 4 培养前后土壤AOA amoA基因DGGE分析 Fig. 4 DGGE analysis based on the archaeal amoA gene before and after the incubation |

|
图 5 DGGE条带中amoA基因序列系统发育树 Fig. 5 Phylogenetic tree based on the archaeal amoA gene in DGGE bands |

|
图 6 培养前后土壤AOA、AOB和NOB相对丰度变化量 Fig. 6 Changes in relative abundance of different AOA, AOB and NOB populations in total pyrosequencing reads before and after the incubation |
3种土壤中Nitrosospira占总AOB类群比例均大于99.99%, 表明Nitrosospira是本研究土壤中最主要的AOB类群(图 6). CK、NPK土壤AOB中的Nitrosomonas和Nitrosospira相对丰度培养前后无显著变化, 而OM土壤的Nitrosomonas相对丰度无显著变化, Nitrosospira相对丰度则从1.22%显著上升至2.18%.
3种土壤的Nitrospira占总NOB类群的比例分别为94.59%、96.25%和96.69%, 表明Nitrospira是本研究土壤中最主要的NOB类群(图 6). NOB群落分析显示, CK、NPK土壤NOB中的Nitrospina、Nitrospira和Nitrobacter相对丰度培养前后无显著性差异; OM土壤的Nitrospina和Nitrobacter相对丰度无显著变化, 而Nitrospira则从1.16%显著上升至2.34%.
微域培养前后AOA和AOB群落组成分析表明:CK和NPK土壤硝化微生物丰度变化较相似, 显示Nitrososphaera属AOA具有活性; 而OM土壤中, Nitrososphaera属AOA、Nitrosospira属AOB和Nitrospira属NOB均具有活性.
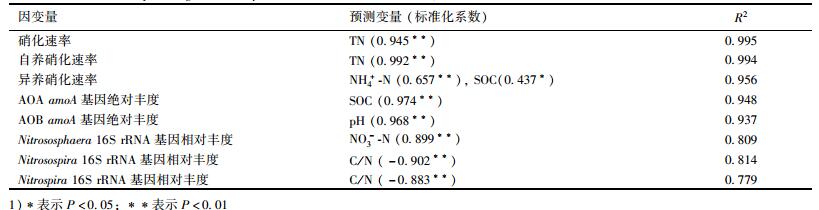
2.5 长期施肥土壤硝化活性及硝化微生物群落变化的环境因子分析逐步回归分析显示(表 2), 土壤总硝化速率显著受TN含量影响, 自养硝化速率也与土壤TN含量显著相关, 异养硝化速率与NH4+-N和SOC含量显著相关; 土壤AOA和AOB amoA基因丰度分别与土壤SOC含量和pH显著相关; 土壤Nitrososphaera属AOA相对丰度与土壤NO3--N含量显著相关; 土壤Nitrosospira属AOB和Nitrospira属NOB相对丰度与C/N显著负相关.
|
|
表 2 土壤硝化活性、硝化微生物丰度与土壤理化性质的逐步回归分析 Table 2 Stepwise regression analysis between nitrification activities, abundance of nitrification oxidizers and soil characteristics |
3 讨论 3.1 长期施肥对酸性旱地土壤硝化活性的影响
土壤硝化作用分为自养硝化和异养硝化, 农业生产活动, 如施肥、灌溉等会改变土壤特性, 从而影响硝化活性.本研究发现施化肥显著提升了土壤的自养硝化速率, 而施有机肥土壤的自养和异养硝化速率均有显著提升, 且施有机肥土壤自养硝化速率增加了6.44倍. Muller等[23]对北爱尔兰长期定位施肥38a的土壤研究显示, 施无机肥和有机肥均会显著提升土壤自养和异养硝化速率, 且硝化速率(尤其是自养硝化)受有机肥的刺激更为明显, 与本研究结果一致. Wang等[24]对我国南部紫色土的研究表明, 相较于不施肥土壤的硝化速率[8.90 mg·(kg·d)-1], 施无机肥和有机肥土壤的硝化速率分别显著上升至23.53 mg·(kg·d)-1和27.81 mg·(kg·d)-1.可见, 施肥给土壤自养硝化作用提供大量底物, 其中施有机肥还会为土壤提供有效碳和其他对改善土壤肥力有益的养分, 提高土壤pH进而促进自养硝化[25].
逐步回归分析表明, 本研究土壤总硝化速率的主要影响因子为TN、NH4+-N; 其中影响自养硝化速率的理化因子为TN, 而异养硝化速率与NH4+-N和SOC含量显著相关. Xue等[26]对酸性茶园土壤(pH 3.71~4.22) 和Zhong等[27]对酸性水稻土壤(pH 5.03~5.77) 的研究也证实了施肥带来的底物含量积累可能是酸性农业土壤硝化作用提升的重要原因.另外, 前人研究发现土壤硝化速率与土壤pH显著正相关[28], 调节酸性土壤pH后, 土壤硝化速率会随土壤pH的显著上升而增加[29].而本研究选用的酸性旱地土壤本底pH极低(pH=3.88), 施化肥导致大量硝化底物流入土壤环境, 但没有加剧土壤酸化, 因此是TN、NH4+-N等硝化底物含量而不是pH,是影响长期定位施肥酸性土壤硝化速率的主控因子.异养硝化是以含氮有机物或有机碳和铵作为碳源和能源氧化氮的过程, 本研究中土壤异养硝化速率的主控因子为NH4+-N和SOC, Wrage等[30]发现高有机碳、低pH的土壤环境均会促进异养硝化, 与本研究结果一致.
3.2 长期施肥对酸性旱地土壤自养硝化微生物的影响本研究发现施肥和不施肥土壤中AOA均有自养硝化活性, 而AOB仅在OM土壤中具有活性, 即施化肥没有影响土壤氨氧化微生物的组成, 而施有机肥改变了土壤活性氨氧化微生物类群.前人运用稳定同位素核酸探针技术发现在中性或者偏碱性(pH 6.90~8.00) 土壤中AOB具有较高氨氧化活性[31, 32]; 但在酸性土壤(pH 4.20~4.47) 中仅检测到AOA具有自养氨氧化活性[15, 33].可见, 相比于AOB, AOA在酸性土壤中能够发挥主导优势的功能活性.分析原因:酸性土壤中氨氧化作用底物浓度较低(氨分子浓度受pH影响, NH3+H+NH4+, pKa=9.25, 即pH越低NH3含量越低), 有利于对氨分子有较高亲和力的AOA的生长[16].有研究表明在NH3含量较低(7.0~87.6 nmol·L-1)的土壤中, AOA是主要的氨氧化执行者; 而在NH3含量较高(6.73~232 μmol·L-1)的土壤中, AOB是主要执行者.其次, AOA具有独特的代谢途径和生理代谢特点以长期适应低pH、低营养等逆境[20, 31~33].本研究发现AOB的丰度与pH显著正相关、与C/N显著负相关.因为长期有机肥施加显著提升土壤pH至近中性, 极大的提升了土壤NH3含量和TN比例, 使得AOB在OM中有明显自养硝化活性; 而CK和NPK土壤呈强酸性, 极大地抑制了AOB活性, 导致在本研究没有检测到AOB活性.
本研究还发现土壤中活跃的AOA类群是Nitrososphaera. Zhang等[34]的研究表明, 长期施无机肥和无机肥-有机肥配施土壤(pH 6.3) 中AOA优势类群为Nitrososphaera, 与本研究结果一致; 而Wang等[15]对长期施肥红壤(pH 3.11~5.55) 的研究认为优势AOA类群却为Nitrosotalea(group 1.1a-associated), 与本研究结果不同.本研究显示酸性土壤中发挥重要作用的AOA类群通常是Nitrosotalea, 且全球尺度[35]和全国尺度[36]上宏基因组研究也发现Nitrososphaera类群相对丰度与土壤pH显著正相关, 而Nitrosotalea类群相对丰度与土壤pH呈显著负相关; 且纯菌株Nitrosotalea devanaterra[37]为嗜酸性菌、Nitrososphaera viennensis EN76和JG1为嗜中性菌[38, 39].但近年来科学家也在酸性农田土壤(pH 4.92) 中发现Nitrososphaera具有功能优势, 且与已报道的嗜中性的Nitrososphaera viennensis EN76高度相似[17], 而笔者在其他酸性土壤中也发现了相似结果(未发表数据).本研究中AOA的丰度与SOC和NO3--N含量显著正相关, 相关研究[13, 40]也发现相似结果.硝酸盐是硝化作用产物, AOA丰度与硝酸盐含量相关, 也证明AOA在本研究酸性土壤硝化过程中发挥了巨大作用.而许多研究还表明AOA具有利用有机底物的潜力或能够受有机底物刺激生长[38, 41], 暗示SOC也会影响AOA在土壤中的角色.可见, group 1.1b在酸性和中性土壤中都能发挥主导作用, 打破了研究者对于AOA群落分布及生态功能的一贯认识, 具体原因还需进一步研究.
另外, 本研究中活跃的AOB类群为Nitrosospira. Nitrosospira是土壤中最为活跃的AOB类群, 在很多土壤中均占有优势[5].本研究还在OM土壤中发现NOB活性明显, 优势类群为Nitrospira, 且与C/N显著负相关.前人对种植冬小麦的长期施肥土壤的研究表明[42], 长期施肥显著增加了土壤Nitrospira的相对丰度, 与本研究结果一致.施有机肥导致大量的硝化底物与养分流入土壤, 进而刺激土壤NOB的增长, TN的增加导致C/N显著降低, 与NOB的增长呈现负相关.
4 结论本研究显示长期施化肥与有机肥均会改变土壤各项理化因子, 土壤TN含量显著刺激酸性旱地土壤自养硝化活性上升, 其中施有机肥刺激更加明显.长期施肥酸性旱地土壤中AOA均有明显活性, 其优势类群为Nitrososphaera, 其丰度与SOC和NO3--N含量显著正相关.仅在施有机肥土壤中检测到AOB(优势类群为Nitrosospira)和NOB(优势类群为Nitrospira)活性, 其中AOB丰度与pH显著正相关、与C/N显著负相关, NOB丰度与C/N显著负相关.
| [1] | Zhang J B, Sun W J, Zhong W H, et al. The substrate is an important factor in controlling the significance of heterotrophic nitrification in acidic forest soils[J]. Soil Biology and Biochemistry, 2014, 76: 143-148. DOI:10.1016/j.soilbio.2014.05.001 |
| [2] |
郭志英, 贾仲君. 中国典型生态系统土壤硝化强度的整合分析[J]. 土壤学报, 2014, 51(6): 1317-1324. Guo Z Y, Jia Z J. Meta-analysis of soil nitrification activity in ecosystems typical of China[J]. Acta Pedologica Sinica, 2014, 51(6): 1317-1324. |
| [3] |
王敬, 程谊, 蔡祖聪, 等. 长期施肥对农田土壤氮素关键转化过程的影响[J]. 土壤学报, 2016, 53(2): 292-304. Wang J, Cheng Y, Cai Z C, et al. Effects of long-term fertilization on key processes of soil nitrogen cycling in agricultural soil: a review[J]. Acta Pedologica Sinica, 2016, 53(2): 292-304. |
| [4] | Nicol G W, Leininger S, Schleper C, et al. The influence of soil pH on the diversity, abundance and transcriptional activity of ammonia oxidizing archaea and bacteria[J]. Environmental Microbiology, 2008, 10(11): 2966-2978. DOI:10.1111/emi.2008.10.issue-11 |
| [5] | Zhong W H, Bian B Y, Gao N, et al. Nitrogen fertilization induced changes in ammonia oxidation are attributable mostly to bacteria rather than archaea in greenhouse-based high N input vegetable soil[J]. Soil Biology and Biochemistry, 2016, 93: 150-159. DOI:10.1016/j.soilbio.2015.11.003 |
| [6] | Wu Y C, Ke X B, Hernández M, et al. Autotrophic growth of bacterial and archaeal ammonia oxidizers in freshwater sediment microcosms incubated at different temperatures[J]. Applied and Environmental Microbiology, 2013, 79(9): 3076-3084. DOI:10.1128/AEM.00061-13 |
| [7] | Chen X P, Zhu Y G, Xia Y, et al. Ammonia-oxidizing archaea: important players in paddy rhizosphere soil?[J]. Environmental Microbiology, 2008, 10(8): 1978-1987. DOI:10.1111/emi.2008.10.issue-8 |
| [8] | Wang B Z, Zhao J, Guo Z Y, et al. Differential contributions of ammonia oxidizers and nitrite oxidizers to nitrification in four paddy soils[J]. The ISME Journal, 2015, 9(5): 1062-1075. DOI:10.1038/ismej.2014.194 |
| [9] |
孟红旗, 刘景, 徐明岗, 等. 长期施肥下我国典型农田耕层土壤的pH演变[J]. 土壤学报, 2013, 50(6): 1109-1116. Meng H Q, Liu J, Xu M G, et al. Evolution of pH in topsoils of typical Chinese croplands under long-term fertilization[J]. Acta Pedologica Sinica, 2013, 50(6): 1109-1116. |
| [10] |
张逸飞, 刘娟娟, 孟磊, 等. 农业利用对海南省天然次生林土壤微生物的影响[J]. 生态学报, 2015, 35(21): 6983-6992. Zhang Y F, Liu J J, Meng L, et al. Agricultural use of natural secondary forests affects soil microorganisms in Hainan Province, China[J]. Acta Ecologica Sinica, 2015, 35(21): 6983-6992. |
| [11] | Xue C, Zhang X, Zhu C, et al. Quantitative and compositional responses of ammonia-oxidizing archaea and bacteria to long-term field fertilization[J]. Scientific Reports, 2016, 6: 28981. DOI:10.1038/srep28981 |
| [12] | Zhou X, Fornara D, Wasson E A, et al. Effects of 44 years of chronic nitrogen fertilization on the soil nitrifying community of permanent grassland[J]. Soil Biology and Biochemistry, 2015, 91: 76-83. DOI:10.1016/j.soilbio.2015.08.031 |
| [13] | Ai C, Liang G Q, Sun J W, et al. Different roles of rhizosphere effect and long-term fertilization in the activity and community structure of ammonia oxidizers in a calcareous fluvo-aquic soil[J]. Soil Biology and Biochemistry, 2013, 57: 30-42. DOI:10.1016/j.soilbio.2012.08.003 |
| [14] | Wu Y C, Lu L, Wang B Z, et al. Long-Term field fertilization significantly alters community structure of ammonia-oxidizing bacteria rather than archaea in a paddy soil[J]. Soil Science Society of America Journal, 2011, 75(4): 1431-1439. DOI:10.2136/sssaj2010.0434 |
| [15] | Wang X L, Han C, Zhang J B, et al. Long-term fertilization effects on active ammonia oxidizers in an acidic upland soil in China[J]. Soil Biology and Biochemistry, 2015, 84: 28-37. DOI:10.1016/j.soilbio.2015.02.013 |
| [16] | Martens-Habbena W, Berube P M, Urakawa H, et al. Ammonia oxidation kinetics determine niche separation of nitrifying Archaea and Bacteria[J]. Nature, 2009, 461(7266): 976-979. DOI:10.1038/nature08465 |
| [17] | Wang B Z, Zheng Y, Huang R, et al. Active ammonia oxidizers in an acidic soil are phylogenetically closely related to neutrophilic archaeon[J]. Applied and Environmental Microbiology, 2014, 80(5): 1684-1691. DOI:10.1128/AEM.03633-13 |
| [18] | He J Z, Shen J P, Zhang L M, et al. Quantitative analyses of the abundance and composition of ammonia-oxidizing bacteria and ammonia-oxidizing archaea of a Chinese upland red soil under long-term fertilization practices[J]. Environmental Microbiology, 2007, 9(9): 2364-2374. DOI:10.1111/emi.2007.9.issue-9 |
| [19] | 鲁如坤. 土壤农业化学分析方法[M]. 北京: 中国农业科技出版社, 2000. |
| [20] | Lu L, Han W Y, Zhang J B, et al. Nitrification of archaeal ammonia oxidizers in acid soils is supported by hydrolysis of urea[J]. The ISME Journal, 2012, 6(10): 1978-1984. DOI:10.1038/ismej.2012.45 |
| [21] | Francis C A, Roberts K J, Beman J M, et al. Ubiquity and diversity of ammonia-oxidizing archaea in water columns and sediments of the ocean[J]. Proceedings of the National Academy of Sciences of the United States of America, 2005, 102(41): 14683-14688. DOI:10.1073/pnas.0506625102 |
| [22] | Rotthauwe J H, Witzel K P, Liesack W. The ammonia monooxygenase structural gene amoA as a functional marker: molecular fine-scale analysis of natural ammonia-oxidizing populations[J]. Applied and Environmental Microbiology, 1997, 63(12): 4704-4712. |
| [23] | Müller C, Laughlin RJ, Christie P, et al. Effects of repeated fertilizer and cattle slurry applications over 38 years on N dynamics in a temperate grassland soil[J]. Soil Biology and Biochemistry, 2011, 43(6): 1362-1371. DOI:10.1016/j.soilbio.2011.03.014 |
| [24] | Wang J, Zhu B, Zhang J B, et al. Mechanisms of soil N dynamics following long-term application of organic fertilizers to subtropical rain-fed purple soil in China[J]. Soil Biology and Biochemistry, 2015, 91: 222-231. DOI:10.1016/j.soilbio.2015.08.039 |
| [25] | Cheng Y, Wang J, Mary B, et al. Soil pH has contrasting effects on gross and net nitrogen mineralizations in adjacent forest and grassland soils in central Alberta, Canada[J]. Soil Biology and Biochemistry, 2013, 57: 848-857. DOI:10.1016/j.soilbio.2012.08.021 |
| [26] | Xue D, Yao H Y, Huang C Y. Microbial biomass, N mineralization and nitrification, enzyme activities, and microbial community diversity in tea orchard soils[J]. Plant and Soil, 2006, 288(1-2): 319-331. DOI:10.1007/s11104-006-9123-2 |
| [27] | Zhong W H, Cai Z C, Zhang H. Effects of long-term application of inorganic fertilizers on biochemical properties of a rice-planting red soil[J]. Pedosphere, 2007, 17(4): 419-428. DOI:10.1016/S1002-0160(07)60051-4 |
| [28] |
鲍俊丹, 石美, 张妹婷, 等. 中国典型土壤硝化作用与土壤性质的关系[J]. 中国农业科学, 2011, 44(7): 1390-1398. Bao J D, Shi M, Zhang M T, et al. Nitrification of main soils in China and its relationship with soil properties[J]. Scientia Agricultura Sinica, 2011, 44(7): 1390-1398. |
| [29] |
张苗苗, 王伯仁, 李冬初, 等. 长期施加氮肥及氧化钙调节对酸性土壤硝化作用及氨氧化微生物的影响[J]. 生态学报, 2015, 35(19): 6362-6370. Zhang M M, Wang B R, Li D C, et al. Effects of long-term N fertilizer application and liming on nitrification and ammonia oxidizers in acidic soils[J]. Acta Ecologica Sinica, 2015, 35(19): 6362-6370. |
| [30] | Wrage N, Velthof G L, van Beusichem M L, et al. Role of nitrifier denitrification in the production of nitrous oxide[J]. Soil Biology and Biochemistry, 2001, 33(12-13): 1723-1732. DOI:10.1016/S0038-0717(01)00096-7 |
| [31] | Jia Z J, Conrad R. Bacteria rather than Archaea dominate microbial ammonia oxidation in an agricultural soil[J]. Environmental Microbiology, 2009, 11(7): 1658-1671. DOI:10.1111/emi.2009.11.issue-7 |
| [32] | Xia W W, Zhang C X, Zeng X W, et al. Autotrophic growth of nitrifying community in an agricultural soil[J]. The ISME Journal, 2011, 5(7): 1226-1236. DOI:10.1038/ismej.2011.5 |
| [33] | Zhang L M, Hu H W, Shen J P, et al. Ammonia-oxidizing archaea have more important role than ammonia-oxidizing bacteria in ammonia oxidation of strongly acidic soils[J]. The ISME Journal, 2012, 6(5): 1032-1045. DOI:10.1038/ismej.2011.168 |
| [34] | Zhang Q, Liang G Q, Myrold D D, et al. Variable responses of ammonia oxidizers across soil particle-size fractions affect nitrification in a long-term fertilizer experiment[J]. Soil Biology and Biochemistry, 2017, 105: 25-36. DOI:10.1016/j.soilbio.2016.11.005 |
| [35] | Gubry-Rangin C, Hai B, Quince C, et al. Niche specialization of terrestrial archaeal ammonia oxidizers[J]. Proceedings of the National Academy of Sciences of the United States of America, 2011, 108(52): 21206-21211. DOI:10.1073/pnas.1109000108 |
| [36] | Hu H W, Zhang L M, Dai Y, et al. pH-dependent distribution of soil ammonia oxidizers across a large geographical scale as revealed by high-throughput pyrosequencing[J]. Journal of Soils and Sediments, 2013, 13(8): 1439-1449. DOI:10.1007/s11368-013-0726-y |
| [37] | Lehtovirta-Morley L E, Stoecker K, Vilcinskas A, et al. Cultivation of an obligate acidophilic ammonia oxidizer from a nitrifying acid soil[J]. Proceedings of the National Academy of Sciences of the United States of America, 2011, 108(38): 15892-15897. DOI:10.1073/pnas.1107196108 |
| [38] | Tourna M, Stieglmeier M, Spang A, et al. Nitrososphaera viennensis, an ammonia oxidizing archaeon from soil[J]. Proceedings of the National Academy of Sciences of the United States of America, 2011, 108(20): 8420-8425. DOI:10.1073/pnas.1013488108 |
| [39] | Kim J G, Jung M Y, Park S J, et al. Cultivation of a highly enriched ammonia-oxidizing archaeon of thaumarchaeotal group Ⅰ. 1b from an agricultural soil[J]. Environmental Microbiology, 2012, 14(6): 1528-1543. DOI:10.1111/j.1462-2920.2012.02740.x |
| [40] |
吕玉, 周龙, 龙光强, 等. 不同氮水平下间作对玉米土壤硝化势和氨氧化微生物数量的影响[J]. 环境科学, 2016, 37(8): 3229-3236. Lü Y, Zhou L, Long G Q, et al. Effect of different nitrogen rates on the nitrification potential and abundance of ammonia-oxidizer in intercropping maize soils[J]. Environmental Science, 2016, 37(8): 3229-3236. |
| [41] | Levi Dč nik-Höfferle Š, Nicol G W, Ausec L, et al. Stimulation of thaumarchaeal ammonia oxidation by ammonia derived from organic nitrogen but not added inorganic nitrogen[J]. FEMS Microbiology Ecology, 2012, 80(1): 114-123. DOI:10.1111/j.1574-6941.2011.01275.x |
| [42] |
李晨华, 张彩霞, 唐立松, 等. 长期施肥土壤微生物群落的剖面变化及其与土壤性质的关系[J]. 微生物学报, 2014, 54(3): 319-329. Li C H, Zhang C X, Tang L S, et al. Effect of long-term fertilizing regime on soil microbial diversity and soil property[J]. Acta Microbiologica Sinica, 2014, 54(3): 319-329. |