 2017, Vol. 38
2017, Vol. 38



2. 华北水利水电大学水务研究院, 郑州 450045
2. Water Affairs Research Institute, North China University of Water Resources and Electric Power, Zhengzhou 450045, China
水体富营养化已成为世界性关注的问题, 在全球范围内30%~40%的湖泊和水库遭受不同程度富营养化的影响[1, 2].水华的发生是在富营养情况下, 藻类的生理学特征与适宜的环境条件和生物因子叠加的生态结果[3].蓝细菌是引起水华的重要组成部分.绝大部分的水华是由浮游微生物(包括蓝细菌门、绿藻门、甲藻门、裸藻门及隐藻门等)引起, 其中蓝细菌水华的发生对环境及人体健康危害最大[4, 5].氮、磷等无机营养物质的过量增加是导致蓝细菌水华暴发的主要原因[6].蓝细菌的最适生长温度一般为25.0~35.0℃, 高于绿藻和硅藻, 这使得蓝细菌在夏秋高温天气更具有竞争力[7~10].同时Zimba等[11]认为水体稳定性、扰动及气候变化等环境因素都会影响蓝细菌的生长及水华的暴发.
北海湖位于北京北海公园内, 是北京城内最大水域之一, 属于城市景观浅水型水体, 由于客流量大而受人为污染风险大, 尤其在旺季(4~11月), 游船项目的开设增加了游客对水体破坏的风险.同时湖水与京城其他水域相连接(上连什刹海, 下接中南海), 北海湖水质受什刹海水质的影响, 同时如果北海水质变差可能会影响中南海的水质.但是关于北海湖水质的数据较少, 并且鲜见报道北海湖微生物的群落结构特征.
采用传统显微镜观察方法研究微生物多样性受多方面条件限制, 如工作量大, 需要观察者具有丰富的微生物观察经验等.但是高通量测序是通过对微生物群落的DNA进行测序, 并与数据库中已有的序列进行比对, 获得微生物的多样性数据.由于该方法不需要人工培养和肉眼鉴定因此工作量小、受人为影响的因素小, 数据量大, 还可能发现新的物种, 因而越来越受研究者的关注.本研究调查北海湖水质随季节的变化特征, 并采用MiSeq高通量测序方法检测北海湖微生物群落结构的季节特征, 分析其与环境因子之间的关系, 旨在通过本研究对北海湖水质和水华风险评价提供参考.
1 材料与方法 1.1 研究区域和样品采集北海湖位于北海公园内(39°55′28.04″N, 116°22′58.83″E), 占公园总面积的54.8%, 湖岸线长3 749 m, 平均水深1.0 m, 最深处达2.5 m.其水源来自颐和园的昆明湖, 流经紫竹院、什刹海, 注入本湖后, 通向中南海及故宫的筒子河, 后经通慧河和海河而流注渤海.在北海湖的出水口处设置采样点(图 1).于2014年9月至2015年8月对该湖水进行采集, 共采集10个时间点的样品(201409、201410、201412、201501、201503、201504、201505、201506、201507和201508;前四个数字为年份, 后两个数字为月份), 按季节分为秋季(201409、201410)、冬季(201412、201501)、春季(201503、201504、201505) 和夏季(201506、201507、201508).每个时间点对采样点均进行3次重复采样取平均值, 每次采集1.0 L水样, 其中0.5 L用于理化性质的测定, 剩余的水样用于水样DNA的提取.

|
图 1 采样点位置示意 Fig. 1 Sampling sites in Beihai Park |
北海湖所属气候为典型的暖温带半湿润大陆性季风气候, 夏季高温多雨, 冬季寒冷干燥, 春、秋短促.汛期一般为每年的6~8月, 封冰期一般为3个月.水文条件(表 1)由北京统计局官方网站(http://www.bjstats.gov.cn/tjsj/)提供, 由表 1可知降雨量季节差异较大, 主要集中在夏季, 夏季降雨量为232.2 mm, 占年降雨量的50.6%;其次为秋季, 降雨量为139.6 mm, 占年降雨量的30.2%;春冬季节降雨相对较小.全年只有在个别月份会出现短暂的大风天气, 不会因风速引起湖面明显波动.
|
|
表 1 天气情况及水文条件 Table 1 Weather and hydrological conditions |

1.2 水质指标的测定
采用便携式水质分析仪(Hach, USA)现场测定水温(T)和溶解氧(DO).其他理化指标(包括化学需氧量COD、生化需要量BOD5、总磷TP和总氮TN)则在水样带回实验室24 h之内完成, 具体测定方法如表 2所示.
|
|
表 2 检测项目方法仪器 Table 2 Methods and instruments |

1.3 DNA的提取和高通量测序
取约300 mL水样, 用0.22 μm的滤膜(47 mm直径的亲水性Durapore表面滤膜, 聚偏氟乙烯材料, Millipore, Ireland, UK)抽滤, 收集一张滤膜的微生物约需1 h.在提取总DNA之前将滤膜保存在-20℃的冰箱内.用无菌剪刀剪碎滤膜后, 采用DNA提取试剂盒(Fast DNA® SPIN Kit for Soil)提取总DNA时, 操作步骤按照试剂盒说明书进行.用1%琼脂糖凝胶电泳检测提取DNA的效果, 并用NanoDrop ND-2000超微量分光光度计(Thermo Scientific, Waltham, MA, USA)定量检测DNA的浓度.将提取得到的DNA保存于-20℃的冰箱中备用.
PCR扩增及其高通量测序采用细菌引物为338F:5′-ACTCCTACGGGAGGCAGCA-3′和806R:5′-GGACTACHVGGGTWTCTAAT-3′.在PCR扩增前, 合成带有barcode(5~8个核苷酸序列)的上述引物用于PCR扩增. PCR扩增采用TransGen AP221-02:TransStart Fastpfu DNA Polymerase, 即全部样本按照正式实验条件进行, 每个样本3个重复, 将同一样本的PCR产物混合后用2%琼脂糖凝胶电泳检测, 使用AxyPrepDNA凝胶回收试剂盒(AXYGEN公司)切胶回收PCR产物, Tris_HCl洗脱; 2%琼脂糖电泳检测.在PCR产物荧光定量后采用Illumina MiSeq PE250/PE300进行高通量测序.测序委托上海美吉生物信息公司进行, 对原始序列进行预处理, 选取高质量的序列进行分析.
1.4 数据分析高通量测序所得的序列通过QIIME(http://qiime.org/tutorials/index.html)进行处理, 主要包括OTU(Operational Taxonomic Units)分类、PCoA(principal co-ordinates analysis)聚类分析、序列的系统发育分析和多性分析.
采用冗余分析(redundancy analysis, RDA)方法, 分析微生物种群与环境因子之间的相关性[12], 其中微生物物种以OTUs代替, 采用R语言进行作图.
2 结果与分析 2.1 水质评估北海湖水温随季节变化明显, 由高到低依次为夏季(27.5℃)、秋季(22.2℃)、春季(19.7℃)、冬季(8.0℃).参考《地表水环境质量标准基本项目标准限值》(GB 3838-2002), 对北海湖水的DO、TN、TP、COD和BOD5进行评价(表 3).
|
|
表 3 不同季节各项理化指标及水质评估(平均值±标准偏差) /mg·L-1 Table 3 Physicochemical indices of different seasons and water quality assessment (Mean±SD)/mg·L-1 |
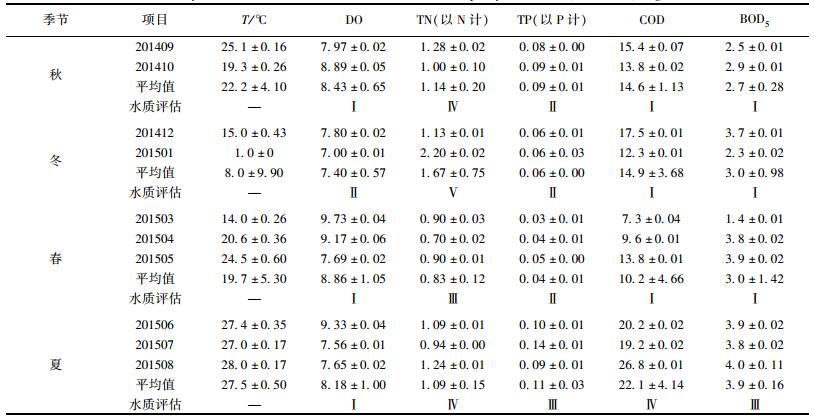
以每季度平均值为标准. DO含量为7.40~8.86 mg·L-1, 其中冬季最低, 符合Ⅱ类水标准, 其他3个季节均为Ⅰ类水. TN含量为0.83~1.67 mg·L-1, 其中冬季TN最高, 为Ⅴ类水, 夏季和秋季含量稍低, 符合Ⅳ类水标准, 春季含量最低, 但也仅达到Ⅲ类水标准. TP含量为0.04~0.11 mg·L-1, 其中夏季含量最高, 符合Ⅲ类水标准, 其他3个季节均为Ⅱ类水. COD为10.2~22.1 mg·L-1, 其中夏季最高, 符合Ⅳ类水标准, 其他3个季节均为Ⅰ类水. BOD5为2.7~3.9 mg·L-1, 夏季含量最高, 符合Ⅲ类水标准, 其他3个季节均为Ⅰ类水.由表 3分析可知, 除夏季COD含量较高外, TN和TP是影响北海湖水质的主要因素.
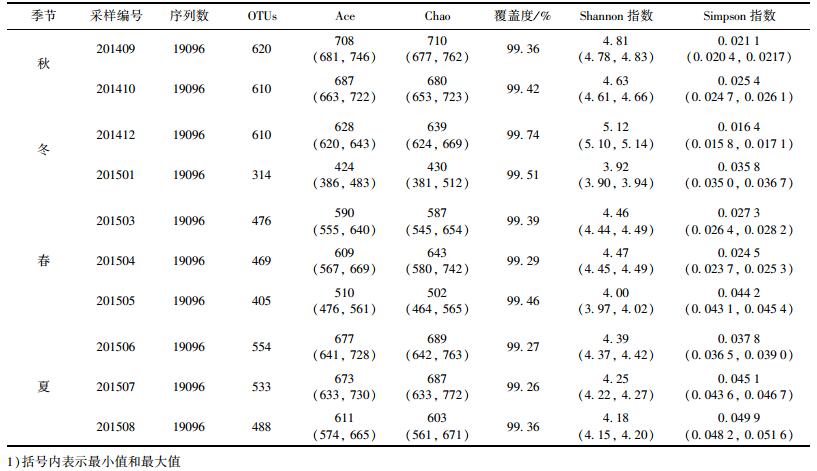
2.2 微生物群落结构如表 4所示, 经过MiSeq测序, 每个样品挑取19 096条优质序列进行分析.本研究将相似度为97%的序列划分为1个OTU, 并进行多样性指数计算.所有样品的覆盖度(coverage)均超过99.0%, 表明细菌测序深度已达到较高水平, 数据可靠.由Ace丰富度指数可知, 夏秋两季群落的丰富度较高, Ace指数高于611, 而冬季和春季样品的Ace均低于628, 冬季201501样品的Ace指数甚至低至424. Chao丰富度指数所表现的趋势和Ace指数相同. Shannon多样性指数季节变化趋势为:夏季<春季<冬季<秋季, Shannon多样性指数常作为一种生物监测的指标来指示水体受有机体污染的状况, 其值越高多样性也越高, 稳定性越强, 水质越好, 其值大于3为轻或无污染, 2~3为β-中污染, 1~2为α-中污染, 0~1为重污染[13, 14].由此判断北海湖全年水质均为轻或无污染. Simpson多样性指数值越大, 群落多样性越低[15], 稳定性越差, 由表 4可知, 其所表现的多样性规律与Shannon多样性指数结果一致.
|
|
表 4 浮游微生物种群丰富度和多样性指数1) Table 4 Summary of the richness and diversity indices of the planktonic microorganism communities |
如图 2所示, 细菌序列主要分布于10个菌门中, 其中放线菌门(Actinobacteria)、变形菌门(Proteobacteria)、蓝细菌门(Cyanobacteria)和拟杆菌门(Bacteroidetes)之和在各个样品中均占79%以上, 但这些菌门的分布有所不同.夏秋样品中(除201506外), 蓝细菌门(Cyanobacteria)是第一优势门, 占全细菌群落的32.7%~43.2%.冬季变形菌门(Proteobacteria)取代蓝细菌门(Cyanobacteria), 成为第一优势门, 占全细菌群落的34.8%~40.4%.春季, 随着气温的升高, 变形菌门(Proteobacteria)的优势作用降低, 放线菌门(Actinobacteria)和拟杆菌门(Bacteroidetes)的相对丰度明显增高, 两者之和占细菌群落的51.5%~64.3%.其他被检测到的菌门包括疣微菌门(Verrucomicrobia)、浮霉菌门(Planctomycetes)、绿菌门(Chlorobi)、绿弯菌门(Chloroflexi)、厚壁菌门(Firmicutes)和梭杆菌门(Fusobacteria).

|
图 2 门分类水平 Fig. 2 Relative abundances of the bacterial phyla |
在属分类水平上, 细菌的群落随季节变化明显.如表 5所示, 真核藻类的叶绿体(Chloroplast)序列在所有样品中的含量均高于2.0%, 其中夏季样品中相对丰度最低, 仅为2.0%~2.3%, 而在冬末(201501) 和初春(201503), 其相对丰度达到15.9%~16.8%.聚球藻属(Synechococcus)和(Order)Subsection I在夏季和秋季含量较高(5.0%~16.9%), 在冬季和春季含量较低(0.3%~4.8%).鞘丝藻属(Leptolyngbya)在夏季和初秋(201409) 含量较高, 分别占全细菌序列的4.3%~15.1%和3.8%, 而在其他样品中的含量仅为0.1%~0.7%. (Order) SubsectionⅢ在夏季含量达到1.1%~2.6%, 初秋(201409) 含量为0.6%, 而在其他样品中的含量低于0.1%.
秋季和冬季变形菌门(Proteobacteria)中属的多样性较高, 其中(Family)Rhodobacteraceae、Albidiferax和GOBB3-C201仅在冬季和春季检测到, 而未在夏季和秋季检测到. Limnohabitans和Polynucleobacter在冬季和春季含量丰富, 而在夏季和秋季含量较低(表 5).
|
|
表 5 不同季节样品中的细菌含量/% Table 5 Bacteria present in the different seasons/% |
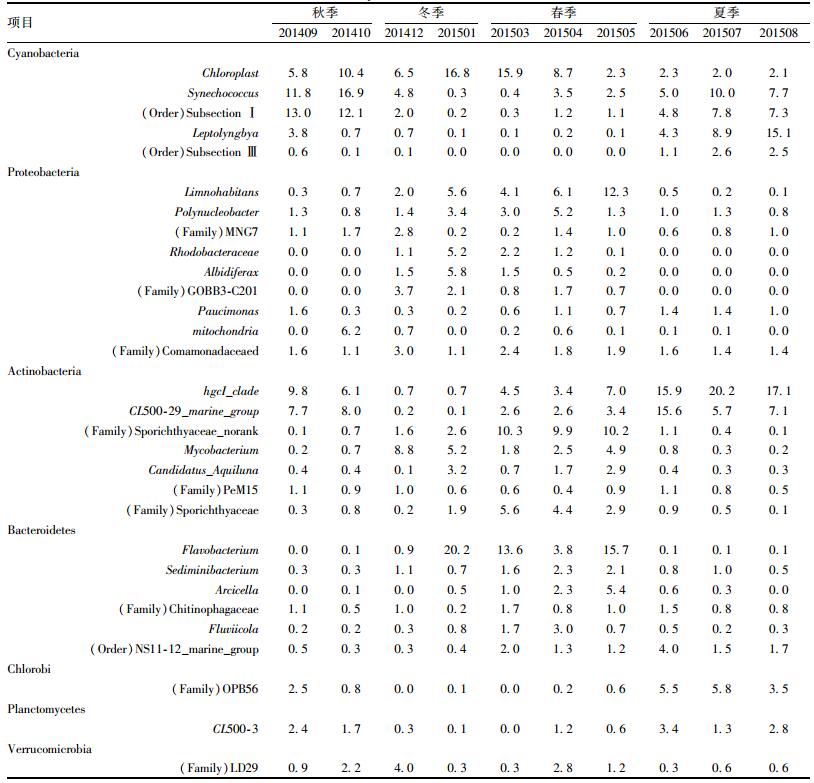
放线菌门(Actinobacteria)中, hgcI_clade的含量由高到底依次为夏季(15.9%~20.2%)、秋季(6.1%~9.8%), 春季(3.4%~7.0%)和冬季(0.7%). CL500-29_marine_group的含量在各个季节的变化趋势与hgcI_clade相似, 含量由高到底依次为夏季(5.7%~15.6%)、秋季(7.7%~8.0%), 春季(2.6%~3.4%)、冬季(0.1%~0.2%).而放线菌门(Actinobacteria)的其他属在春季含量较高.
拟杆菌门(Bacteroidetes)中的黄杆菌属(Flavobacterium)在冬季和春季节含量较高, 分别为0.9%~20.2%和3.8%~15.7%, 但是在夏季和秋季的样品中含量仅为0.1%.
本研究中检测到其他属的存在, 如在绿藻门(Chlorobi)中的OPB56和浮霉菌门(Planctomycetes)中的CL500-3仅在夏季和秋季含量丰富, 而在冬季和春季含量较低.
2.3 聚类分析采用PCoA分析衡量样本间群落物种组成的相似度(图 3), 结果表明四个季节分布在不同位置, 因此不同月份水体间物种还是存在较大差异, 从图 3可以看出, 结合主成分PC1以及主成分PC2进行分析, 算出PC1和PC2对物种分布的总解释量为76.64%.夏秋季节微生物群落结构相似度高, 冬春季节微生物群落结构相似度高, 除冬季外, 同一季节内的样品相似度高.因此可将夏秋数据分为一组, 春冬季节分为一组.进而将这两个分组的Ace和Chao指数进行方差分析, 结果也表明夏秋季节和春冬季节多样性存在显著性差异(P=0.03).

|
图 3 PCoA分析 Fig. 3 PCoA plot |
采用RDA分析方法分析北海湖水体理化性质对细菌群落结构的影响.由图 4可知, 两个坐标轴RDA1与RDA2对细菌群落结构分布的总解释度为67.11%. T对北海湖水体细菌群落结构影响最大, 其次是TP和TN. T和TN对夏季(除201508外)样品分别呈正相关和负相关. TP对秋季样品与夏季(201508) 呈正相关.

|
图 4 北海湖微生物与环境因子的RDA分析 Fig. 4 RDA of microbiological and environmental factors |
北海湖水质符合景观水标准, 夏秋季节水质劣于春冬季节.北海湖水体中TN是影响水质的关键因素, 尤其在冬季(201501) 其含量高达2.20 mg·L-1, 这可能由于冬季水温低, 水体中微生物的活性较低, 导致TN的降解效果较差[16, 17]. TP是衡量水体受污染程度的一个重要指标, 当水体中磷含量过高时, 水质变坏, 因此TP是水质中不可或缺的指标[18].依据文献[19]规定湖泊营养类型, 北海湖水体秋冬季节为富营养型, 春季为中营养型, 夏季为重营养型.究其原因, 首先上游污染严重的水体流入湖内是导致水体理化指标超标的重要原因之一, 什刹海作为北海湖上游, 2007~2013年TN、TP含量较高, 陶亮等[20]在2013年调查研究也曾发现, “六海”TN含量较高主要原因是上游河道将TN含量明显较高的水体输入导致[21].其次降雨径流可能使外源性污染物流入湖内造成污染, 张青青[22]指出降雨径流污染和内源释放污染已经成为城市景观水体氮磷等营养盐的主要来源, 苟婷等[23]也曾指出强降雨会将土壤中积累的营养盐流入水体.降雨也会引起湖泊水体水文和水动力学特征的变化, 从而影响营养盐的变化[24].采样期间, 夏秋季节降雨量分别占年降雨量的50.6%和30.2%, 明显高于其他两季, 致使夏秋季节的水质劣于春冬.另外旅游污染也可能引起水体污染, 北海公园为北京名胜公园之一, 旅游旺期(4~11月)每日接待游客至少约2万人, 游客往湖内投食、丢弃废物相比其他季节更为严重[25].对此, 要进一步加大对上游水源的监督与控制; 加强对雨污合流排水管的改造, 减轻降雨径流对湖泊的危害; 同时公园管理人员要加强管理力度, 对游客丢弃垃圾、投食喂鱼等不文明现象及时制止, 甚至可以在适当的时候控制游客数量.
3.2 北海湖细菌群落结构分析细菌多样性指数即Shannon多样性是反映多样性综合性指标, 既包括物种丰富度又包括均匀度[26].袁聪等[27]、张华等[28]发现夏季Shannon多样性指数高于冬季, 但本研究结果却与之相反, 本研究中夏季的Ace和Chao丰富度指数均高于冬季, 由于夏季样品中的某些优势物种的丰度随温度降低而降低, 低丰度物种随温度降低而升高, 导致冬季样品中群落的均匀度高于夏季, 进而使得北海湖冬季微生物多样性整体高于夏季.
蓝细菌门(Cyanobacteria)、拟杆菌门(Bacteroidetes)、放线菌门(Actinobacteria)和变形菌门(Proteobacteria)是北海湖水体中的主要菌门, 这与大量研究湖水细菌群落结构的文献相似[29~32], 但不同菌门的分布呈现明显的季节特征, 且细菌的群落结构与环境因子的关系密切.
由RDA分析可知, T对北海湖微生物群落结构影响最大, 且夏季和秋季样品的微生物群落分布与T和TP呈正相关性. Brogueira等[33]和Cameiro等[34]在Tagus河口研究发现T也是影响微生物群落结构(主要为浮游藻类)的主要因子.这是由于夏秋季节水温偏高, 营养物质丰富的环境中微生物生长较旺盛, 进而导致不同环境出现较大的微生物群落差异[35].北海湖夏秋季节蓝细菌(Cyanobacteria)的相对丰度较高, 原因是蓝细菌(Cyanobacteria)嗜高温的特性致使其在夏秋季节大量繁殖所致[36, 37]. Zarina等[38]在巴基斯坦淡水栖息地也曾发现蓝细菌(Cyanobacteria)在高温下生长旺盛.一般而言, TP的含量越高蓝细菌(Cyanobacteria)的相对丰度也越高, 和本研究发现的夏季和秋季TP含量较高, 蓝细菌(Cyanobacteria)的相对丰度也较高的结果一致[39]. T和TP的变化导致蓝细菌(Cyanobacteria)丰度发生变化, 进一步影响了北海湖整体微生物群落结构的季节性变化.水质分析结果表明夏季水体中COD增加, 主要原因也和蓝细菌(Cyanobacteria)大量繁殖而进行更多的光合作用有关.而COD的变化使得夏季样品微生物群落结构发生变化.聚球藻属(Synechoccus)是超微蓝细菌的一种, 是初级生产力的主要参与者和贡献者, 在生态系统的光合作用、碳循环及食物链中具有重要作用并且很难通过形态学观察检测到, 已在太湖、滇池和其他富营养化的淡水湖泊中发现该菌属大量存在[40~44].北海湖夏秋季节水体中的聚球藻属(Synechoccus)含量丰富, 因此应对其水体加强监测和管理, 避免蓝细菌水华的暴发.本研究中检测Subsection Ⅰ和Subsection Ⅲ中未分类的(Order)Subsection Ⅰ和(Order)Subsection Ⅲ(表 5)与数据库中已有的序列相似度低, 标志北海湖中有特异性的微生物存在.与蓝细菌(Cyanobacteria)的分布不同, 真核微生物(藻类)在夏季的相对丰度最低, 而冬季的相对丰度较高, 研究者针对微生物(藻类)随温度的分布特征发现, 硅藻与温度呈现负相关, 在10℃相对丰度达到最大[45].
变形菌门(Proteobacteria)通常是水环境中主要的原核生物, 包含了光营养性、化能自养型和化能异养型细菌, 这类细菌在水环境中普遍存在, 相关报道指出在湿地、养殖废水和深海沉积物等环境中以第一优势门存在[46~49], 也是本研究冬季样品中的优势门. Limnohabitans作为主要的淡水浮游生物, TN和TP对其生长繁殖有重要的作用[50].本研究结果发现Limnohabitans在夏秋季节含量丰富, 这和本次研究中发现夏秋季节水体中TN、TP含量高的现象相符.这也间接反映出夏秋季节水质较差, 与水质评价结果相一致, 因此在夏秋季节更要加强水质监测避免发生水体富营养化.文献[51~53]分别在天津海河、巴西Paraguaçú河和黄海海岸淡水泉中发现Limnohabitans大量存在.
放线菌门(Actinobacteria)和拟杆菌门(Bacteroidetes)是水体中主要的微生物, 承担水体的基本功能, 是本研究中春季样品的优势门.放线菌门(Actinobacteria)中的hgcI_clade属在秋季和夏季样品中含量较高(6.1%~20.2%).这与王鹏等[54]研究赣江南昌段丰水期细菌群落特征时hgcI_clade属含量最高(16.39%)结果较为相似. hgcI_clade具有较强的吸收碳水化合物和富氮化合物的遗传能力, 能在较低的DO含量下生存, 还能利用低温水中的低浓度溶解有机碳[55, 56].拟杆菌门(Bacteroidetes)中的黄杆菌属(Flavobacterium)在冬末、春初和春末3个时间段含量最高(13.6%~20.2%).卢思丹等[57]在沣河微生物群落中发现它以优势属出现; 秦煊[58]调查鄱阳湖微生物群落结构时也曾发现它是优势属.黄杆菌属(Flavobacterium)主要分布在淡水湖泊和土壤中靠分解有机物生存, 具有广泛的溶藻能力, 使得水质在一定程度得以提高[59, 60], 这与本研究观察到此段时间水质优于夏秋季节的现象相符合.另外, 黄杆菌属(Flavobacterium)的某些菌种具有致病作用, 集中分布在淡水河水、江河、土壤沉积物和鱼的发病组织中, 主要表现在对鱼的致病作用[61].同时它还可以引起医院内感染, 尤其是婴幼儿更容易感染[62].因此, 对于该菌属也不容忽视, 要加强防护意识.
4 结论总体而言, 北海湖水在夏季和秋季的水质偏差, TN和TP是影响水质的主要因素.环境因子和微生物的关系密切, TN和TP有助于微生物的生长, 会引起微生物的大量繁殖, 微生物通过光合作用, 固定空气中的CO2, 从而导致水体COD增高, 引起水质的恶性循环. T是影响微生物群落结构的关键因素:夏季和秋季水温较高, 微生物的多样性最高, 蓝细菌的分布尤为广泛, 其中聚球藻属(Synechococcus)大量存在, 可能会引起水华的暴发, 因此应保持对北海湖水质和微生物的检测, 预防水华的发生.
致谢: 对北京市北海公园及相关工作人员在现场采样给予的帮助表示感谢!| [1] |
王圣瑞, 倪兆奎, 席海燕. 我国湖泊富营养化治理历程及策略[J]. 环境保护, 2016, 44(18): 15-19. Wang S R, Ni Z K, Xi H Y. Management process and strategy of lake eutrophication in China[J]. Environmental Protection, 2016, 44(18): 15-19. |
| [2] |
袁琦. 上海水源水中微生物群落结构的分子生态学研究[D]. 上海: 华东师范大学, 2011. Yuan Q. Molecular ecology research on microbial community in the water of waterhead area in Shanghai[D]. Shanghai: East China Normal University, 2011. |
| [3] |
张艳会, 李伟峰, 陈求稳. 太湖水华程度及其生态环境因子的时空分布特征[J]. 生态学报, 2016, 36(14): 4337-4345. Zhang Y H, Li W F, Chen Q W. Spatial-temporal variance of the intensity of algal bloom and related environmental and ecological factors in Lake Taihu[J]. Acta Ecologica Sinica, 2016, 36(14): 4337-4345. |
| [4] | 莫婉湫, 李策, 刘玉, 等. 认识"水华"[J]. 湿地科学与管理, 2009, 5(1): 62-63. |
| [5] | 谢平. 论蓝藻水华的发生机制——从生物进化、生物地球化学和生态学视点[M]. 北京: 科学出版社, 2007. |
| [6] | WHO, UNESCO, UNEP. Toxic cyanobacteria in water[M]. London: St. Edmundsbury Press, 1999. |
| [7] |
胡传林, 万成炎, 吴生桂, 等. 蓝藻水华的成因及其生态控制进展[J]. 长江流域资源与环境, 2010, 19(12): 1471-1477. Hu C L, Wan C Y, Wu S G, et al. Progress in causes and ecological control of cyanobacterial bloom[J]. Resources and Environment in the Yangtze Basin, 2010, 19(12): 1471-1477. |
| [8] |
郑维发, 曾昭琪. 淡水蓝藻的高温适应[J]. 湖泊科学, 1994, 6(4): 356-363. Zheng W F, Zeng Z Q. High temperature adaptation of fresh water cyanobacterium[J]. Journal of Lake Sciences, 1994, 6(4): 356-363. DOI:10.18307/1994.0409 |
| [9] | Robarts R D, Zohary T. Temperature effects on photosynthetic capacity, respiration, and growth rates of bloom-forming cyanobacteria[J]. New Zealand Journal of Marine and Freshwater Research, 1987, 21(3): 391-399. DOI:10.1080/00288330.1987.9516235 |
| [10] | Yamaguchi M, Ogawa T, Muramoto K, et al. Effects of culture conditions on the expression level of lectin in Microcystis aeruginosa (freshwater cyanobacterium)[J]. Fisheries Science, 2000, 66(4): 665-669. DOI:10.1046/j.1444-2906.2000.00106.x |
| [11] | Zimba P V, Khoo L, Gaunt P S, et al. Confirmation of catfish, Ictalurus punctatus(Rafinesque), mortality from Microcystis toxins[J]. Journal of Fish Diseases, 2001, 24(1): 41-47. DOI:10.1046/j.1365-2761.2001.00273.x |
| [12] | Xiao L J, Wang T, Hu R, et al. Succession of phytoplankton functional groups regulated by monsoonal hydrology in a large canyon-shaped reservoir[J]. Water Research, 2011, 45(16): 5099-5109. DOI:10.1016/j.watres.2011.07.012 |
| [13] |
余海霞, 来勇, 赵佳佳, 等. 城市河道浮游植物群落多样性指数对水质的指示作用[J]. 化学与生物工程, 2016, 33(9): 32-36. Yu H H, Lai Y, Zhao J J, et al. Water quality indication of phytoplankton community diversity index in urban rivers[J]. Chemistry & Bioengineering, 2016, 33(9): 32-36. |
| [14] |
朱爱民, 胡菊香, 李嗣新, 等. 三峡水库长江干流及其支流枯水期浮游植物多样性与水质[J]. 湖泊科学, 2013, 25(3): 378-385. Zhu A M, Hu J X, Li S X, et al. Phytoplankton diversity and water quality in the main stream and tributaries of Three Gorges Reservoir region of Yangtze River during dry season[J]. Journal of Lake Sciences, 2013, 25(3): 378-385. DOI:10.18307/2013.0311 |
| [15] |
马永红, 曾燏, 任丽萍, 等. 嘉陵江四川段藻类植物群落结构及水质评价[J]. 应用生态学报, 2012, 23(9): 2573-2579. Ma Y H, Zeng Y, Ren L P, et al. community structure of phycophyta and evaluation of water quality in Sichuan section of Jialing River[J]. Chinese Journal of Applied Ecology, 2012, 23(9): 2573-2579. |
| [16] |
聂泽宇, 梁新强, 邢波, 等. 基于氮磷比解析太湖苕溪水体营养现状及应对策略[J]. 生态学报, 2012, 32(1): 48-55. Nie Z Y, Liang X Q, Xing B, et al. The current water trophic status in Tiaoxi River of Taihu Lake watershed and corresponding coping strategy based on N/P ratio analysis[J]. Acta Ecologica Sinica, 2012, 32(1): 48-55. |
| [17] | 刘婷婷. 嘉陵江水体中碳、氮、磷季节变化及其输出[D]. 重庆: 西南大学, 2009. |
| [18] |
王爱爱, 冯佳, 谢树莲. 汾河中下游浮游藻类群落特征及水质分析[J]. 环境科学, 2014, 35(3): 915-923. Wang A A, Feng J, Xie S L. Phytoplankton community structure and assessment of water quality in the middle and lower reaches of Fenhe River[J]. Environmental Science, 2014, 35(3): 915-923. |
| [19] | 金相灿, 屠清瑛. 湖泊富营养化调查规范[M]. 第二版. 北京: 中国环境科学出版社, 1990. |
| [20] | 陶亮, 杨菊, 孙峰. "六海"富营养化态势及对策研究[J]. 北京水务, 2014(4): 33-35. |
| [21] |
周坤朋, 刘阳春, 王崇臣. 北京什刹海区域水体富营养化时空演变特征分析[J]. 环境化学, 2016, 35(4): 703-712. Zhou K P, Liu Y C, Wang C C. Analysis on temporal-spatial variation of eutrophication in Shichahai area, Beijing[J]. Environmental Chemistry, 2016, 35(4): 703-712. DOI:10.7524/j.issn.0254-6108.2016.04.2015093001 |
| [22] |
张青青. 城市景观水体氮磷污染负荷来源解析[D]. 西安: 西安建筑科技大学, 2015. Zhang Q Q. The nitrogen and phosphorus pollution of the landscape water[D]. Xi'an: Xi'an University of Architecture and Technology, 2015. |
| [23] |
苟婷, 马千里, 许振成, 等. 贝江浮游藻类群落特征及富营养化风险分析[J]. 环境科学, 2015, 36(3): 946-954. Gou T, Ma Q L, Xu Z C, et al. Phytoplankton community structure and eutrophication risk assessment of Beijing River[J]. Environmental Science, 2015, 36(3): 946-954. |
| [24] | O'Farrell I, Izaguirre I, Chaparro G, et al. Water level as the main driver of the alternation between a free-floating plant and a phytoplankton dominated state: a long-term study in a floodplain lake[J]. Aquatic Sciences, 2011, 73(2): 275-287. DOI:10.1007/s00027-010-0175-2 |
| [25] |
王迎春, 任天志, 王立刚, 等. 北京市水体富营养化的成因与治理对策分析[J]. 中国农业资源与区划, 2005, 26(3): 40-44. Wang Y C, Ren T Z, Wang L G, et al. Analysis on reasons and harness counter measures for nutritive water in Beijing[J]. Chinese Journal of Agricultural Resources and Regional Planning, 2005, 26(3): 40-44. |
| [26] |
任丽娟, 何聃, 邢鹏, 等. 湖泊水体细菌多样性及其生态功能研究进展[J]. 生物多样性, 2013, 21(4): 421-432. Ren L J, He D, Xing P, et al. Bacterial diversity and ecological function in lake water bodies[J]. Biodiversity Science, 2013, 21(4): 421-432. |
| [27] |
袁聪, 陶诗雨, 张莹莹, 等. 安康水库表层浮游藻类群落结构及其与环境因子的关系[J]. 应用生态学报, 2015, 26(7): 2167-2176. Yuan C, Tao S Y, Zhang Y Y, et al. community structure of phytoplankton and its relationships with environmental factors in surface water of Ankang Reservoir, Northwest China[J]. Chinese Journal of Applied Ecology, 2015, 26(7): 2167-2176. |
| [28] |
张华, 胡鸿钧, 晁爱敏, 等. 浙江紧水滩水库浮游植物群落结构季节变化特征[J]. 生态学报, 2013, 33(3): 944-956. Zhang H, Hu H J, Chao A M, et al. Seasonal changes of phytoplankton community structure in Jinshuitan Reservoir, Zhejiang, China[J]. Acta Ecologica Sinica, 2013, 33(3): 944-956. |
| [29] |
陈楠. 太湖沉积物微生物群落组成与物质循环及藻华爆发的相关性[D]. 北京: 中国农业大学, 2015. Chen N. Relationship between microbial community structure and nutrient cycles and algal bloom in Taihu Lake[D]. Beijing: China Agricultural University, 2015. |
| [30] | Parfenova V V, Gladkikh A S, Belykh O I. comparative analysis of biodiversity in the planktonic and biofilm bacterial communities in Lake Baikal[J]. Microbiology, 2013, 82(1): 91-101. DOI:10.1134/S0026261713010128 |
| [31] | Kaluzhnaya O V, Itskovich V B. Distinctive features of the microbial diversity and the polyketide synthase genes spectrum in the community of the endemic Baikal sponge Swartschewskia papyracea[J]. Russian Journal of Genetics, 2016, 52(1): 38-48. DOI:10.1134/S1022795416010099 |
| [32] | Bai F, Zeng S Y, Li D, et al. Analysis of the bacterial community by 454 pyrosequencing in two freshwater lakes: Yangcheng Lake and Taihu Lake in China[J]. Advanced Materials Research, 2013, 788: 315-320. DOI:10.4028/www.scientific.net/AMR.788 |
| [33] | Brogueira M J, do Rosário Rosario M, Cabeçadas G. Phytoplankton community structure defined by key environmental variables in Tagus estuary, Portugal[J]. Marine Environmental Research, 2007, 64(5): 616-628. DOI:10.1016/j.marenvres.2007.06.007 |
| [34] | Gameiro C, Cartaxana P, Brotas V. Environmental drivers of phytoplankton distribution and composition in Tagus Estuary, Portugal[J]. Estuarine, Coastal and Shelf Science, 2007, 75(1-2): 21-34. DOI:10.1016/j.ecss.2007.05.014 |
| [35] |
孙寓姣, 陈程, 丁爱中, 等. 官厅水库水质特征及水体微生物多样性的响应[J]. 中国环境科学, 2015, 35(5): 1547-1553. Sun Y J, Chen C, Ding A Z, et al. The corresponding of microbial diversity on water quality and environmental variables of Guanting Reservoir[J]. China Environmental Science, 2015, 35(5): 1547-1553. |
| [36] |
田昌. 洪泽湖浮游植物种群结构变化的水环境驱动因子分析[D]. 济南: 山东大学, 2015. Tian C. Variation of phytoplankton community and its driving factors in Hongze Lake[D]. Ji'nan: Shandong University, 2015. |
| [37] | Liu Z H, Huang S B, Sun G P, et al. Phylogenetic diversity, composition and distribution of bacterioplankton community in the Dongjiang River, China[J]. FEMS Microbiology Ecology, 2012, 80(1): 30-44. DOI:10.1111/j.1574-6941.2011.01268.x |
| [38] | Zarina A, Shameel M, Naz S. Distribution of freshwater blue-green algae (Cyanophyta) in northeastern Pakistan[J]. International Journal on Algae, 2010, 12(3): 257-270. DOI:10.1615/InterJAlgae.v12.i3 |
| [39] | Hu A Y, Yang X Y, Chen N W, et al. Response of bacterial communities to environmental changes in a mesoscale subtropical watershed, Southeast China[J]. Science of the Total Environment, 2014, 472: 746-756. DOI:10.1016/j.scitotenv.2013.11.097 |
| [40] | Raven J A. The twelfth Tansley lecture. Small is beautiful: the picophytoplankton[J]. Functional Ecology, 1998, 12(4): 503-513. DOI:10.1046/j.1365-2435.1998.00233.x |
| [41] |
李胜男, 周建, 魏利军, 等. 淡水超微型浮游植物多样性及其研究方法[J]. 生态学杂志, 2015, 34(4): 1174-1182. Li S N, Zhou J, Wei L J, et al. A review of diversity of freshwater picophytoplankton and its related research methods[J]. Chinese Journal of Ecology, 2015, 34(4): 1174-1182. |
| [42] | 冯丽君. 太湖蓝藻多样性及群落结构时空动态研究[D]. 上海: 华东理工大学, 2016. |
| [43] | 李艳晖. 滇池、洱海超微浮游蓝藻、绿藻细胞学和多样性研究[D]. 武汉: 中国科学院水生生物研究所, 2009. |
| [44] | Bailey-Watts A E, Bindloss M E, Belcher J H. Freshwater primary production by a blue-green alga of bacterial size[J]. Nature, 1968, 220(5174): 1344-1345. DOI:10.1038/2201344a0 |
| [45] | 曹方意. 太湖流域浮游藻类特征及对水生态功能分区的指示意义[D]. 南京: 南京工业大学, 2015. |
| [46] | Spring S, Schulze R, Overmann J, et al. Identification and characterization of ecologically significant prokaryotes in the sediment of freshwater lakes: molecular and cultivation studies[J]. FEMS Microbiology Reviews, 2000, 24(5): 573-590. DOI:10.1111/j.1574-6976.2000.tb00559.x |
| [47] |
杜瑞芳, 李靖宇, 赵吉. 乌梁素海湖滨湿地细菌群落结构多样性[J]. 微生物学报, 2014, 54(10): 1116-1128. Du R F, Li J Y, Zhao J. Bacterial diversity in littoral wetland of Wuliangsuhai lake[J]. Acta Microbiologica Sinica, 2014, 54(10): 1116-1128. |
| [48] |
张健. 海参循环水养殖系统建立及曝气生物滤池细菌多样性分析[D]. 哈尔滨: 哈尔滨工业大学, 2013. Zhang J. Establishment of recirculating aquaculture system of sea cucumber and analysis of bacterial diversity in biological aerated filter[D]. Harbin: Harbin Institute of Technology, 2013. |
| [49] |
陈丽蓉. 北极深海沉积物中微生物的多样性研究[D]. 杭州: 浙江理工大学, 2012. Chen L R. The investigation on microbial diversity of Arctic deep sea sediments[D]. Hangzhou: Zhejiang Sci-Tech University, 2012. |
| [50] | Salcher M M. Same same but different: ecological niche partitioning of planktonic freshwater prokaryotes[J]. Journal of Limnology, 2013, 73(S1): 74-87. |
| [51] | Ma L L, Mao G N, Liu J, et al. Spatial-temporal changes of bacterioplankton community along an exhorheic river[J]. Frontiers in Microbiology, 2016, 7: 250. |
| [52] | Lopes F A C, Catão E C P, Santana R H, et al. Microbial community profile and water quality in a protected area of the Caatinga biome[J]. PLoS One, 2016, 11(2): e0148296. DOI:10.1371/journal.pone.0148296 |
| [53] | Ye Q, Liu J, Du J Z, et al. Bacterial diversity in submarine groundwater along the coasts of the Yellow Sea[J]. Frontiers in Microbiology, 2016, 6: 1519. |
| [54] |
王鹏, 陈波, 李传琼, 等. 赣江南昌段丰水期细菌群落特征[J]. 中国环境科学, 2016, 36(8): 2453-2462. Wang P, Chen B, Li C Q, et al. Bacterial communities in Nanchang section of the Ganjiang River in wet seaon[J]. China Environmental Science, 2016, 36(8): 2453-2462. |
| [55] | Ghylin T W, Garcia S L, Moya F, et al. comparative single-cell genomics reveals potential ecological niches for the freshwater acI Actinobacteria lineage[J]. The ISME Journal, 2014, 8(12): 2503-2516. DOI:10.1038/ismej.2014.135 |
| [56] | Liu J W, Fu B B, Yang H M, et al. Phylogenetic shifts of bacterioplankton community composition along the Pearl Estuary: the potential impact of hypoxia and nutrients[J]. Frontiers in Microbiology, 2015, 6: 64. |
| [57] |
卢思丹, 孙寓姣, 赵轩, 等. 降水对沣河水质和水体微生物的影响[J]. 环境科学, 2016, 37(7): 2563-2569. Lu S D, Sun Y J, Zhao X, et al. Impact of precipitation on Fenghe River water and aquatic microorganisms[J]. Environmental Science, 2016, 37(7): 2563-2569. |
| [58] |
秦煊. 基于高通量DNA测序的鄱阳湖微生物生态研究[D]. 南昌: 南昌大学, 2015. Qin X. Study on the microbial ecology of Poyang Lake based on next-generation sequencing[D]. Nanchang: Nanchang University, 2015. |
| [59] | da Costa M S, Albuquerque L, Nobre M F, et al. The identification of polar lipids in prokaryotes[J]. Methods in Microbiology, 2011, 38: 165-181. DOI:10.1016/B978-0-12-387730-7.00007-3 |
| [60] |
吴刚, 席宇, 赵以军. 溶藻细菌研究的最新进展[J]. 环境科学研究, 2002, 15(5): 43-46. Wu G, Xi Y, Zhao Y J. The latest development of research on algae-lysing bacteria[J]. Research of Environmental Sciences, 2002, 15(5): 43-46. |
| [61] |
高远. 发现于茅台地区的黄杆菌Flavobacterium maotaiensis sp. nov. 和Flavobacterium renhuaiensis sp. nov. 的分类学研究[D]. 北京: 北京化工大学, 2014. Gao Y. Flavobacterium maotaiensis sp. nov. and Flavobacterium renhuaiensis sp. nov., isolated from Maotai Town[D]. Beijing: Beijing University of Chemical Technology, 2014. |
| [62] | 尚建中, 马光远, 巫善明, 等. 黄杆菌属感染[J]. 中国实用内科杂志, 1995, 15(11): 695-696. |