 2017, Vol. 38
2017, Vol. 38



2. 山东省胶东调水局, 济南 250100;
3. 深圳职业技术学院建筑与环境工程学院, 深圳 518055;
4. 深圳市水质检测中心, 深圳 518055;
5. 清华大学深圳研究生院, 深圳 518055;
6. 深圳市国家气候观象台, 深圳 518040
2. Shandong Province Jiaodong Water Transfer Bureau, Ji'nan 250100, China;
3. School of Architectural and Environmental Engineering, Shenzhen Polytechnic, Shenzhen 518055, China;
4. Shenzhen Water Quality Testing Center, Shenzhen 518055, China;
5. Graduate School at Shenzhen, Tsinghua University, Shenzhen 518055, China;
6. Shenzhen National Climate Observatory, Shenzhen 518040, China
深圳人均年水资源拥有量不足200 m3, 约为全国平均水平的1/12, 属严重缺水城市[1], 全市近90%的饮用水需从境外调取.深圳的供水水源体系主要由境外引水工程以及本地调蓄水库群组成.境外来水经市内中小型水库群存储调节后, 再转供各水厂.截至2015年12月, 深圳共建有93宗供水型水库, 这些中小型供水水库群构成了深圳供水框架的重要组成, 其水质对饮用水安全起到关键作用.
石岩水库位于宝安区石岩街道和光明区公明街道境内, 属茅洲河流域范围.水库集雨面积44 km2, 总库容9 950万m3, 50%可供水量3 400万m3, 是深圳市城市供水网络骨干工程之一, 供水量居全市各水库第2位, 保障了宝安西部地区600多万人口的生产和生活用水.由于流域人口稠密, 虽然实行封闭式管理, 但仍有生活、农业和工业废水流入石岩水库, 使其成为深圳12座主要供水水库中污染最严重的水库, 季节性藻类暴发风险不断加剧[2, 3].目前关于石岩水库的研究较少且多以水质方面研究为主[4, 5], 反映石岩水库藻类年际演替特征的研究相对缺乏.国内外学者对藻华的研究多关注春夏时期的各类天然湖泊[6~8], 对饮用水水源型水库关注偏少, 对氮磷等营养盐关注较多[9~11], 对水质和气象因子的综合影响研究较少.
叶绿素a是藻类进行光合作用的重要色素, 反映了水体的初级生产力[12, 13].通过检测ρ(Chla)估计水体藻类生物量已经成为国际上常用的技术手段[14~16].课题组显微镜计数数据显示蓝藻、硅藻和绿藻是石岩水库的主要藻类, 3种藻类细胞丰度之和占总细胞丰度的95%以上.因此掌握蓝藻、绿藻和硅藻的演替规律是研究石岩水库藻类群落动态的关键.
对2013年4月至2014年6月石岩水库蓝藻、绿藻和硅藻的ρ(Chla)及其主要相关环境因子进行了连续监测, 研究了藻类时空分布特征和演变规律, 分析了水质因子和气象因子对藻类演替的驱动作用, 以期为有效认识藻类生长和群落演替规律, 探索藻华机制, 识别和控制藻类风险, 保障水生态安全提供科学依据, 对深圳乃至珠三角地区同类型的中小型供水水库也具有借鉴意义.
1 材料与方法 1.1 采样点的布设石岩水库由南向北共布设8个采样点, 其中1号采样点靠近铁岗水库进水口, 2号采样点附近有工农业废水进入, 3号采样点靠近河流, 4、5、6号采样点均为库心活水区, 7、8号采样点靠近水厂取水口.采样点具体分布情况见图 1.每个采样点由上到下分3~5层采样, 表层是水面下10 cm左右水层, 透光层是所测透明度深度对应水层, 温跃层是水体1 m内水温(WT)变化超过1℃水层, 活性层是溶解氧(DO)为2 mg·L-1水层, 底层是底泥上方约10 cm水层, 其中表层、透光层和底层始终纳入监测范围, 活性层和温跃层视WT和DO分布状况而定, 出现即监测.

|
图 1 石岩水库采样点布设示意 Fig. 1 Sampling sites of Shiyan Reservoir |
采用ZPY-1型采水器分层采样并分别保存, 水样采集后2 h内运回实验室, 4℃保存, 当天完成ρ(Chla)的测定, 总有机碳(TOC)、高锰酸盐指数(COD)、总氮(TP)、总磷(TN)、氨氮(NH4+-N)、硝酸盐氮(NO3--N)、铁(Fe)和硅酸盐(Silicate)等2 d内完成检测. Chla采用调制荧光仪(德国WALZ Phyto-PAM)测定, 并定期与文献[17]中的丙酮萃取分光光度法校正.水深、WT、pH、DO、浊度(Turbidity)采用多参数水质测定仪(美国YSI 6600V2型)现场测定; 透明度(SD)采用塞氏盘现场测定. COD采用酸性高锰酸钾法测定, TP采用钼酸铵分光光度法测定, Silicate采用硅钼蓝分光光度法测定, TN、NH4+-N、NO3--N采用流动分析仪(法国Alliance-Futura)测定, TOC采用总有机碳分析仪(GE-Siever 5310C)测定.气温、气压、湿度、降雨量、蒸发量和日照时数等气象数据由深圳国家气候观测台提供.
1.3 时期划分根据气象特征将全年划分为5个时期:前汛期为2013、2014年4~6月, 后汛期为2013年7~9月, 高温无雨期为2013年10~11月, 冬季枯水期为2013年12月~2014年2月, 温度跃升期为2014年3月.
1.4 数据处理环境因子以及藻类Chla的时间或空间差异采用单因素方差分析(One-way ANOVA)法分析, 时间和空间因素对环境因子和藻类Chla的交互影响采用两因素方差分析(Two-way ANOVA)法分析, 二者均在SPSS 20上实现; 藻类Chla与环境因子间的双变量关系采用Pearson相关分析在SPSS 20上进行, 藻类Chla与环境因子间的综合相关关系采用CCA在XLSTAT2010上进行.藻类叶绿素a平面分布图是根据各采样点的ρ(TChla)在相应气候期内的多月平均值, 通过克里金插值法, 在ArcMap10.2上生成.
2 结果与讨论 2.1 环境因子动态石岩水库2013年4月~2014年6月环境因子动态见表 1.单因素方差分析结果显示, 时间上WT、SD、COD、TOC、Silicate、NO3--N、TN和Fe等理化因子季节差异显著(P < 0.01), 其中WT、TOC、Silicate、NO3--N、TN和Fe表现为前汛期>后汛期>高温无雨期>温度跃升期>冬季枯水期的规律, SD则表现出相反的规律. DO、pH、NH4+-N和TP等理化因子季节性差异不明显.空间上DO、SD、COD、TOC、TN、NH4+-N和Fe等理化因子随采样点不同而差异显著(P < 0.01), 其中COD、TOC、TN、NH4+-N和Fe由南向北逐渐降低, DO和SD由北向南逐渐升高, 7、8号采样点是石岩水库水质最好的区域. WT、pH、Silicate、TP和NO3--N等理化因子空间差异不明显. TN是石岩水库首要污染物质, 根据地表水环境评价方法的评价结果, 在监测时期内所有采样点, TN年均2.13 mg·L-1, 属地表水劣Ⅴ类水质, 最大值为3.10 mg·L-1, 出现在2013年4月的2号点表层, 最小值为1.58 mg·L-1. DO、COD、TP等指标达到地表水环境质量Ⅱ~Ⅲ类标准.
|
|
表 1 环境因子年际变化规律 Table 1 Annual trends of environment factors for Shiyan Reservoir |
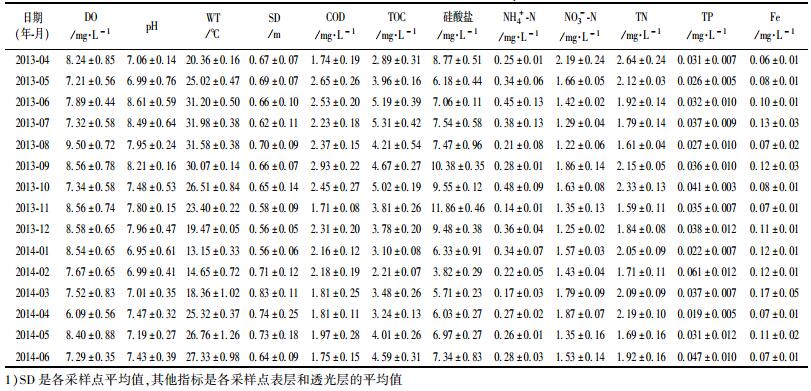
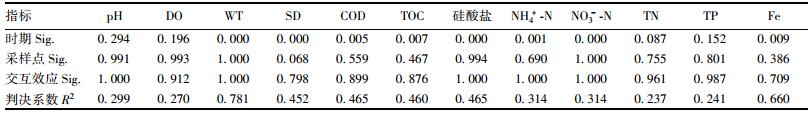
两因素方差分析(表 2)显示, 各环境因子的变异能被时期和采样点及两者交互效应较好地解释(R2为23.7%~78.1%).时期是影响环境因子的最主要因素, 对WT、SD、COD、TOC、Silicate、NH4+-N、NO3--N和Fe均有显著影响(P < 0.01), 与TN也存在一定相关性(P=0.087), 对pH、DO和TP影响较小.采样点对SD有较为显著的影响(P=0.068), 对其他环境因子影响很小(P>0.3).时期和采样点的交互作用对Fe和SD有微弱影响, 对其他环境因子影响甚微(P>0.8).综合来看, 石岩水库环境因子的主要影响因素是时期, 环境因子的时间异质性明显高于空间异质性.
|
|
表 2 时期和采样点对环境因子的交互影响两因素方差分析 Table 2 Two-way ANOVA of period and sampling sites to environmental factors |
2.2 叶绿素a时空演变规律
图 2反映了石岩水库各水层总绿素a浓度年际变化规律.石岩水库藻类ρ(TChla)均值为52.03 μg·L-1, 最高为123.29 μg·L-1, 出现在2013年6月1号点透光层, 最低为9.59 μg·L-1, 出现在2013年12月周采样的2号点底表层.石岩水库藻类季节性暴发现象明显:藻类4月开始复苏, 5~7月达到全年最高水平, 其中7月ρ(TChla)均值为89.04 μg·L-1, 到8~10月藻类浓度略微降低, 11月至来年2月降至低浓度水平, 2013年12月ρ(TChla)均值为16.72 μg·L-1, 为全年最低峰, 2014年3~4月随着温度升高, 藻类再次复苏, 5~6月进入高藻期.与去年同期相比, 2014年5~6月藻类浓度降低25%左右.这是因为石岩水库截污排海工程减少了流域藻类和污染物入库量, 起到了明显改善水质的作用.藻类垂直方向上分布规律与水深存在明显相关性, ρ(TChla)分布规律是透光层>表层>底层, 水深越深, 规律越明显.当水深为3~5 m时, 藻类呈现透光层略大于表层大于底层的分布规律; 当水深为5~8 m时, 底层ρ(TChla)均值迅速降低, 与表层和透光层差距拉大; 当水深为8~15 m时, 底层ρ(TChla)均值仅为透光层的3/5左右.石岩水库藻类叶绿素a在垂直方向上的空间异质性主要是由光照和水温的分层引起的.表层和透光层光照充足, 水温较高则ρ(TChla)高.而底层光照微弱, 水温较低则ρ(TChla)明显偏低.由于风力作用引发的水体垂直方向上的交换, 以及具有伪空胞的藻类会通过浮力调节机制从表层移至深水层, 没有发生底层和表层水体中ρ(TChla)差距悬殊的现象.

|
各组柱由左至右采样点编号为1~8 图 2 石岩水库总绿素a浓度年际变化规律 Fig. 2 Annual trends in total chlorophyll a concentration for Shiyan Reservoir |
石岩水库ρ(TChla)均值虽然低于太湖、巢湖和滇池等污染较重的自然湖泊, 但作为饮用水水源水库, 从ρ(Chla)均值这一指标来看, 石岩水库已经达到了轻度富营养化状态[18, 19].
图 3反映了石岩水库蓝藻、绿藻和硅藻的年际演变规律, 蓝藻ρ(Chla)均值为28.39 μg·L-1, 最高为76.30 μg·L-1, 出现在2013年7月的3号点透光层, 最低为4.54 μg·L-1, 出现在2014年2月的1号点底层; 月均最高为2013年7月56.02 μg·L-1, 最低为2013年12月5.76 μg·L-1.硅藻年ρ(Chla)为15.02 μg·L-1, 最高为46.09 μg·L-1, 出现在2013年6月的1号点透光层, 最低为3.16 μg·L-1, 出现在2013年8月下旬的6号点底层; 月均最高为2013年5月27.35 μg·L-1, 最低为2013年11月7.37 μg·L-1.绿藻ρ(Chla)为8.62 μg·L-1, 最高为26.21 μg·L-1, 出现在2013年5月的8号点透光层, 最低为0.77 μg·L-1, 2013年8月多个采样点底层均有出现; 月均最高为2013年5月15.33 μg·L-1, 最低为2013年8月1.37 μg·L-1.

|
各组柱由左至右采样点编号为1~8 图 3 石岩水库蓝藻、绿藻、硅藻叶绿素a浓度年际变化规律 Fig. 3 Annual trends in chlorophyll a concentrations of cyanophyta, Bacillariophyta, and Chlorophyta for Shiyan Reservoir |
图 4反映了藻类种群结构变化规律, 蓝藻优势明显, 全年占比为53.22%;硅藻次之, 全年占比为30.09%, 在冬春季节占优势; 绿藻全年占比为13.72%, 始终处于较低水平.蓝藻2013年4月~12月上旬一直是优势藻:2013年4月蓝藻比例为48.77%, 之后快速上升, 7月达到最高峰(69.79%), 至11月一直维持高浓度水平, 之后比例迅速下降, 至12月下旬失去优势地位, 2014年4月上旬比例维持在31.33%~36.86%, 4月下旬迅速蹿升, 超越硅藻成为优势藻, 6月比例达到73.13%.硅藻的生长趋势与蓝藻相反:2013年4月硅藻比例为45.67%, 与蓝藻相当, 随后迅速下降, 直至9月比例再次上升, 12月下旬超越蓝藻成为优势藻, 至2014年4月上旬, 比例维持在42.25%~50.05%.绿藻比例为5.57%~23.33%, 全年处于较低水平, 季节性变化不明显.与去年同期相比, 2014年5~6月石岩水库藻类群落结构变化明显:蓝藻优势度明显上升, 硅藻优势度下降, 藻类多样性降低.藻类种群结构是水体生态多样性和稳定性的反映[20~22], 总体来看, 石岩水库藻类结构较为单一, 多样性较差, 藻类生态系统稳定性差, 存在一定蓝藻水华风险.汛期蓝藻产生的藻毒素对饮用水安全造成一定威胁; 冬春季节硅藻的大量繁殖易堵塞给水厂滤池, 增加反冲洗次数, 相伴产生的土臭素和二甲基异莰醇等臭味物质增加了给水厂的处理成本.

|
图 4 石岩水库藻类种群结构变化 Fig. 4 Community structure of phytoplankton in Shiyan Reservoir |

|
图 5 不同气候期内石岩水库Chla浓度平面分布 Fig. 5 Chlorophyll a charts for different climatic periods in Shiyan Reservoir |
石岩水库ρ(Chla)由南到北逐渐降低, 整体呈现中南库区高北部库区低的特点.汛期和高温无雨期多数采样点ρ(Chla)偏高, 存在较高藻华风险. 1、2、3号采样点所处的南半库区ρ(TChla)为60.35 μg·L-1, 是石岩水库ρ(Chla)最高的区域.其中1号采样点是铁岗水库来水进入石岩水库的位置, ρ(Chla)为14.06~123.29 μg·L-1, 均值45.69 μg·L-1.铁岗水库来水中含有高浓度的藻类和营养盐, 使得该区域ρ(Chla)基数大, 藻类生长速率高. 2、3号采样点受径流污染较重, 水体营养状态较高, Chla浓度分别为16.61~109.69 μg·L-1和15.34~119.62 μg·L-1.南半库区在汛期和高温无雨期均存在较高的藻华风险. 4、5、6号采样点位于库心活水区, 该区域水体交换频繁, 水体流速达15~30 cm·s-1, 一定程度上抑制了藻类的快速繁殖, ρ(Chla)顺次降低, 汛期存在一定藻华风险. 7、8号采样点近水厂取水口, 远离径流污染, 是石岩水库水质最优处, 藻类总ρ(Chla)分别为8.96~89.69 μg·L-1和5.11~77.89 μg·L-1, 在汛期存在较高藻华风险.总体看来, 石岩水库面积较小, 形状较为规则, 库内水体交换充分, 各采样点空间异质性较低.径流污染和工农业污染源的汇入是石岩水库藻类叶绿素a空间异质性的主要原因, 因此加强水源保护, 从源头上减少外源污染物的进入是降低藻华风险的根本途径.
2.4 叶绿素a与环境因子关联性分析 2.4.1 叶绿素a与环境因子Pearson相关分析表 3反映了汛期和旱期藻类叶绿素a与水质因子的相关关系, 不同时期不同藻类的主要相关因子差异较大.蓝藻汛期的主要相关因子有WT、TOC、TN:TP、TN、pH、NO3--N、SD等, 其中TN:TP、TN、SD、NO3--N为负相关; 旱期主要相关因子有TOC、WT、NO3--N、DO、TP等, 其中TP为负相关.硅藻汛期主要相关因子有NO3--N、Silicate、TOC、COD、WT、TN:TP, 其中TN:TP、NO3--N为负相关; 旱期主要相关因子有Silicate、DO、NO3--N、TP等, 其中NO3--N、TP为负相关.绿藻汛期主要相关因子有WT、TOC、COD、pH、SD、NO3--N、TN:TP等, 其中NO3--N、TN:TP为负相关; 旱期主要相关因子有Silicate、WT、DO、NO3--N、TP等, 其中NO3--N、TP为负相关.
|
|
表 3 藻类叶绿素a与水质因子Pearson相关系数1) Table 3 Pearson's correlation coefficients between chlorophyll a and meteorological factors |
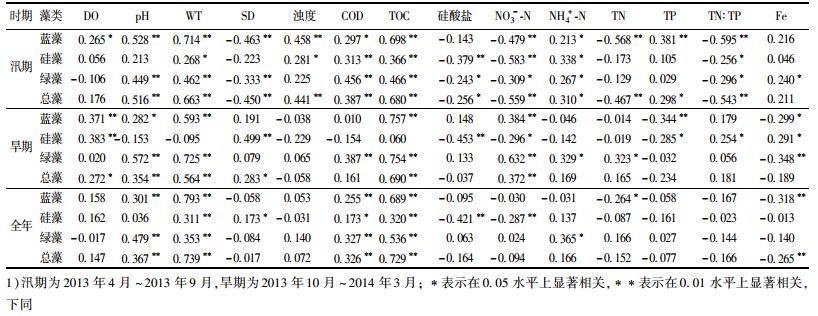
汛期水温是蓝藻的首要影响因子, 相关系数高达0.714, 对硅藻则是第6位的影响因子, 相关系数为0.268.旱期水温是蓝藻次要相关影响因子, 相关系数高达0.593, 对硅藻影响更加微弱, 相关系数为-0.095.水温是影响亚热带湖库藻类生长的首要因素, 这一点与文献[23~25]的研究结论一致.深圳地处亚热带地区, 监测期内水温在14.08~32.33℃范围波动.汛期5~9月水温达29℃以上, 与蓝藻的最适生长温度重合[26~28], 随着温度的升高, 蓝藻在种间竞争中获得优势, 抑制了硅藻的生长.旱期水温为14.08~23.57℃, 适宜硅藻的生长, 硅藻取代蓝藻成为优势藻. 2013年1月~2014年2月随着温度的降低, 蓝藻急速衰减, 硅藻数量反而迅速增加. TOC和COD代表水体有机物含量, 二者与藻类存在较高的相关性.这是因水体绝大部分有机物都会以TOC的形式被检测出来, 而藻类本身就是有机物, 是TOC的重要组成部分.汛期透明度、浊度与藻类尤其是蓝藻呈显著负相关, 这说明汛期藻类的增加, 增加了水体的浑浊度, 降低了太阳辐射在水体的透射性, 限制藻类光合作用.硝酸盐氮、TN:TP、TN等与3种藻类成负相关关系, 石岩水库TN平均浓度为2.03 mg·L-1, 超过地表水Ⅴ类标准, TP平均浓度0.03 mg·L-1, 达到地表水Ⅱ~Ⅲ类标准, 总氮超标明显, 氮磷比为67左右.汛期随着TP增加藻类ρ(Chla)升高, TN的增加反而引起藻类ρ(Chla)降低, 这说明石岩水库高浓度的氮含量能够满足藻类生长需求, 而TP浓度偏低, 是藻类生长的限制性因子.
2.4.2 叶绿素a与气象因子Pearson相关分析同一天各采样点的气象因子数值相同, 数据特征与水质因子不同, 因此单独分析其与藻类Chla的相关性.由表 4可知.不同藻类在同一时期以及同一藻类在不同时期的主要相关因子均存在明显差异.
|
|
表 4 藻类叶绿素a与气象因子Pearson相关系数 Table 4 Pearson's correlation coefficients between chlorophyll a and environmental factors |
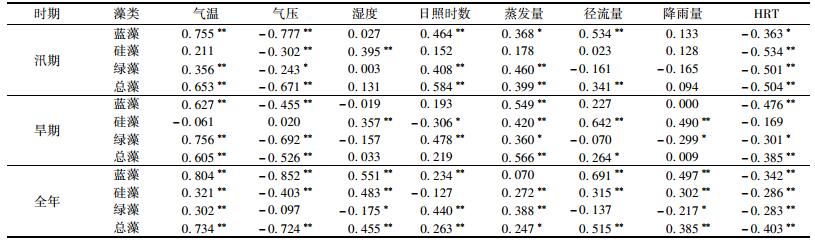
蓝藻与气象因子相关性较高, 在汛期的主要相关因子为气压>气温>径流量, 在旱期的主要相关因子为气温>蒸发量>HRT.硅藻与气象因子相关性一般, 在汛期的主要相关因子为HTR>湿度>气压, 在旱期的主要相关因子为径流量>降雨量>蒸发量.绿藻在汛期与气象因子相关性低, 主要相关因子为HTR>蒸发量>日照时数, 在旱期与气象因子显著相关, 主要相关因子为气温>气压>日照时数.
气压与藻类Chla成显著负相关, 尤其是汛期与蓝藻相关系数达到-0.777, 是蓝藻的首要相关因子.气压与藻类间没有因果关系, 但这种高度相关性说明持续的低压天气容易暴发蓝藻水华.气温与藻类Chla的相关规律与水温基本一致, 汛期与蓝藻显著相关, 旱期与绿藻显著相关, 与硅藻无显著相关性.降雨和径流量是影响藻类Chla的重要因子, 汛期径流量、降雨量与蓝藻相关系数分别为0.534和0.133, 旱期径流量、降雨量与硅藻相关系数分别为0.642和0.490.地表和植被着生的藻类被降雨冲刷下来后随地表径流进入水库.绿藻与径流量的相关性很小, 这可能是因为流域内地表中生长的藻类主要是蓝藻和硅藻. HRT与3种藻类均存在一定负相关性, 说明水力停留时间短, 水体流速加大, 更新速度加快, 可以抑制藻类的生长.因此优化水利调度, 加快水体流动性, 降低HRT, 是抑制藻类水华的有效途径.藻类Chla与蒸发量和日照时数呈一定相关性, 与湿度无明显相关.
2.4.3 叶绿素a与环境因子典范对应分析将汛期和旱期的水质和气象因子与藻类ρ(Chla)进行典范对应分析, 研究了二者综合作用对藻类生长的影响, 结果如图 6所示.汛期蓝藻、绿藻和硅藻分别位于第一、二、三象限, 旱汛期蓝藻、绿藻和硅藻分别位于第一、三、四象限, 说明影响3种藻类的环境因子差异较大.汛期F1和F2的特征值分别为0.015和0.012, 解释率分别为55.71%和44.29%;旱期F1和F2的特征值分别为0.067和0.011, 解释率分别为86.87%和13.13%.

|
汛期2013年4月~2013年9月, 样本数为80;旱期2013年10月~2014年3月, 样本数为71;蓝色、绿色、紫色和黑色圆点代表蓝藻、绿藻、硅藻和总藻, 深蓝色点表示采样点, 箭头线表示环境因子 图 6 石岩水库绿素a浓度随时间变化 Fig. 6 Canonical correspondence analysis of environmental factors and phytoplankton of Shiyan Reservoir |
汛期影响藻类生长和分布的首要因子为径流量, 尤其是蓝藻与径流量高度正相关.石岩水库周边人口稠密, 入库支流包括石岩河、王家庄河、麻布水、后桥河、上屋河、运牛坑水、白坑窝水等, 将流域内的生活废水和工农业废水带入水库, 增加了营养盐水平.同时地表和植物表面附着的藻类经降雨冲洗后随地表径流进入水库, 藻类生物量相应增加.因此加强水库污水截排和内源污染的清除, 限制库区农业和养殖业污染源是解决石岩水库水体富营养化的首要途径.气温、水温、气压、日照时数是影响藻类生长的第二类影响因素, 因此要加强对汛期高温天气藻类监测, 预防暴发局部藻类水华. TOC、COD、TN、氨氮、TP等有机物和营养盐是藻类第三大类影响因子. DO、透明度、HRT和蒸发量与藻类的相关性相对较小, 硅酸盐、降雨量和Fe等与藻类没有明显相关性.
旱期影响藻类生长和分布的首要因子为硅酸盐, 水温和气温的影响效应位居第二位.朱广伟等[29]发现太湖硅藻在气温为16~26℃快速生长, 当气温超过26℃时, 硅藻的生物量开始下降.石岩水库冬季表层水温13~24℃, 水温更适宜硅藻的生长, 硅藻取代蓝藻成为优势藻.旱期降水稀少, 随地表径流进入水库的硅酸盐显著减少, 而硅酸盐正是硅藻生长所需的必要营养盐, 因此硅酸盐与藻类的相关关系越发显著.
TOC和COD代表水中的有机物含量, 与藻类相关性较高, 是第三类影响因素. NO3--N、NH4+-N、TN和TP代表营养盐水平, 是藻类生长的必须因子, NO3--N、NH4+-N、TN与藻类显著相关, TP与藻类相关性较低. Fe与藻类相关性显著, 因为Fe是藻类光合作用和新陈代谢的必要元素, 对藻类生物量和生长速率均有重要影响[30, 31].气压、日照时数、降雨量、径流量、湿度等气象因子与藻类呈一定相关性, 但显著性较汛期有所降低. DO、HRT、蒸发量和TN:TP等与藻类没有明显相关性.
3 结论(1) 石岩水库藻类ρ(TChla)均值为52.03 μg·L-1, 处于轻度富营养化状态, 蓝藻、硅藻和绿藻ρ(Chla)均值分别为28.39、15.02、8.62 μg·L-1. 2013年12月下旬至2014年4月上旬硅藻为优势藻(42%~49%), 其他时间蓝藻优势明显(44%~78%), 绿藻全年处于低浓度水平(6%~23%).
(2) 石岩水库ρ(Chla)时间上变化规律为前汛期>后汛期>高温无雨期>温度跃升期>秋冬储水期, 空间上自南向北逐渐降低.汛期整个库区和高温无雨期中南库区存在蓝藻水华风险, 冬春季节暴发的硅藻虽未达到藻华程度, 但易堵塞给水厂滤池, 妨害供水安全.
(3) 相关分析和CCA分析显示:蓝藻、硅藻、绿藻的关键环境影响因子差异较大, 水温是3种藻类的首要影响因子, 前汛期径流带来的有机物和营养盐是蓝藻暴发的重要诱因.氮磷比与3种藻类负相关, 磷可能是石岩水库藻类生长的限制性因子.蓝藻主要相关因子还包括TOC、TN、pH、透明度、浊度和硝酸盐氮等, 硅藻主要相关因子还包括硅酸盐、TOC、硝酸盐氮和COD等, 绿藻主要相关因子还包括TOC、COD、pH、透明度和硝酸盐氮等.
| [1] |
邱国玉, 张清涛. 快速城市化过程中深圳的水资源与水环境问题[J]. 河海大学学报(自然科学版), 2010, 38(6): 629-633. Qiu G Y, Zhang Q T. Issues of water resources and environment in Shenzhen with rapid urbanization[J]. Journal of Hohai University (Natural Sciences), 2010, 38(6): 629-633. |
| [2] |
温美丽, 方国祥, 陈朝辉, 等. 深圳石岩水库污染状况及综合防治对策[J]. 热带地理, 2009, 29(1): 5-10. Wen M L, Fang G X, Chen C H, et al. Pollution and prevention measures of Shiyan reservoir in Shenzhen city[J]. Tropical Geography, 2009, 29(1): 5-10. |
| [3] |
彭溢, 谢林伸. 深圳市石岩水库富营养化主控因素分析及潜在风险预测[J]. 安全与环境学报, 2012, 12(4): 141-146. Peng Y, Xie L S. Analysis of the main controlling eutrophication factors and prediction of the potential risk of Shiyan Reservoir, Shenzhen[J]. Journal of Safety and Environment, 2012, 12(4): 141-146. |
| [4] | 张琳. 深圳石岩水库清淤过程水质变化研究[D]. 广州: 中国科学院广州地球化学研究所, 2007. |
| [5] |
林高松, 黄晓英, 李娟. 基于投影寻踪法的水库富营养化评价-以深圳市铁岗水库与石岩水库为例[J]. 热带地理, 2010, 30(2): 130-134. Lin G S, Huang X Y, Li J. Eutrophication assessment of reservoirs based on project pursuit method: a case study of Tiegang Reservoir and SHIYAN Reservoir in Shenzhen[J]. Tropical Geography, 2010, 30(2): 130-134. |
| [6] | Tran D V, Gabric A, Cropp R. Interannual variability in chlorophyll-a on the southern Queensland continental shelf and its relationship to ENSO[J]. Journal of Sea Research, 2015, 106: 27-38. DOI:10.1016/j.seares.2015.09.007 |
| [7] | Lira G A S T, Moura A N, Vilar M C P, et al. Vertical and temporal variation in phytoplankton assemblages correlated with environmental conditions in the Mundaú reservoir, semi-arid northeastern[J]. Brazilian Journal of Biology, 2014, 74(S3): S93-S102. |
| [8] | Ma J R, Brookes J D, Qin B Q, et al. Environmental factors controlling colony formation in blooms of the cyanobacteria Microcystis spp. in Lake Taihu, China[J]. Harmful Algae, 2014, 31: 136-142. DOI:10.1016/j.hal.2013.10.016 |
| [9] | Daggett C T, Saros J E, Lafrancois B M, et al. Effects of increased concentrations of inorganic nitrogen and dissolved organic matter on phytoplankton in boreal lakes with differing nutrient limitation patterns[J]. Aquatic Sciences, 2015, 77(3): 511-521. DOI:10.1007/s00027-015-0396-5 |
| [10] | Zhang X F, Mei X Y, Gulati R D, et al. Effects of N and P enrichment on competition between phytoplankton and benthic algae in shallow lakes: a mesocosm study[J]. Environmental Science and Pollution Research, 2015, 22(6): 4418-4424. DOI:10.1007/s11356-014-3680-3 |
| [11] | Zhang W Y, Lou I, Ung W K, et al. Spatio-temporal variations of phytoplankton structure and water quality in the eutrophic freshwater reservoir of Macau[J]. Desalination and Water Treatment, 2015, 55(8): 2237-2252. DOI:10.1080/19443994.2014.930933 |
| [12] |
孙德勇, 周晓宇, 李云梅, 等. 基于光学分类的太湖水体叶绿素a浓度高光谱遥感[J]. 环境科学, 2013, 34(8): 3302-3309. Sun D Y, Zhou X Y, Li Y M, et al. Hyperspectral remote sensing of chlorophyll a concentrations in the Lake Taihu, based on water optical classification[J]. Environmental Science, 2013, 34(8): 3302-3309. |
| [13] |
杨斌, 钟秋平, 张晨晓, 等. 钦州湾叶绿素a和初级生产力时空变化及其影响因素[J]. 环境科学学报, 2015, 35(5): 1333-1340. Yang B, Zhong Q P, Zhang C X, et al. Spatio-temporal variations of chlorophyll a and primary productivity and its influence factors in Qinzhou Bay[J]. Acta Scientiae Circumstantiae, 2015, 35(5): 1333-1340. |
| [14] |
张磊, 蔚建军, 付莉, 等. 三峡库区回水区营养盐和叶绿素a的时空变化及其相互关系[J]. 环境科学, 2015, 36(6): 2061-2069. Zhang L, Wei J J, Fu L, et al. Temporal and spatial variation of nutrients and chlorophyll a, and their relationship in Pengxi river backwater area, three gorges reservoir[J]. Environmental Science, 2015, 36(6): 2061-2069. |
| [15] |
王斌, 马健, 王银亚, 等. 天山天池夏季叶绿素a的分布及富营养化特征研究[J]. 环境科学, 2015, 36(7): 2465-2471. Wang B, Ma J, Wang Y Y, et al. Distribution of chlorophyll-a and eutrophication state in Tianchi Lake of Tianshan mountains in summer[J]. Environmental Science, 2015, 36(7): 2465-2471. |
| [16] | Nazeer M, Nichol J E. Development and application of a remote sensing-based Chlorophyll-a concentration prediction model for complex coastal waters of Hong Kong[J]. Journal of Hydrology, 2016, 532: 80-89. DOI:10.1016/j.jhydrol.2015.11.037 |
| [17] | 国家环境保护总局. 水和废水监测分析方法[M]. 第四版. 北京: 中国环境科学出版社, 2002: 722-723. |
| [18] |
陈藜藜, 金腊华. 湖库富营养化的改进型模糊综合评价方法研究[J]. 中国环境科学, 2014, 34(12): 3223-3229. Chen L L, Jin L H. An improved fuzzy comprehensive evaluation method of eutrophication for lakes and reservoirs[J]. China Environmental Science, 2014, 34(12): 3223-3229. |
| [19] |
陈林, 许其功, 李铁松, 等. 模糊物元识别模型在巢湖水体富营养化评价中的应用研究[J]. 环境工程学报, 2010, 4(4): 729-736. Chen L, Xu Q G, Li T S, et al. Fuzzy pattern recognition model to water trophic grade in Chaohu Lake[J]. Chinese Journal of Environmental Engineering, 2010, 4(4): 729-736. |
| [20] |
马沛明, 施练东, 张俊芳, 等. 浙江汤浦水库浮游植物季节演替及其影响因子分析[J]. 环境科学, 2016, 37(12): 4560-4569. Ma P M, Shi L D, Zhang J F, et al. Succession of phytoplankton assemblages and its influencing factors in Tangpu reservoir, Zhejiang Province[J]. Environmental Science, 2016, 37(12): 4560-4569. |
| [21] |
黄廷林, 曾明正, 邱晓鹏, 等. 温带季节性分层水库浮游植物功能类群的时空演替[J]. 中国环境科学, 2016, 36(4): 1157-1166. Huang T L, Zeng M Z, Qiu X P, et al. Phytoplankton functional groups and their spatial and temporal distribution characteristics in a temperate seasonally stratified reservoir[J]. China Environmental Science, 2016, 36(4): 1157-1166. |
| [22] |
王宇飞, 赵秀兰, 何丙辉, 等. 汉丰湖夏季浮游植物群落与环境因子的典范对应分析[J]. 环境科学, 2015, 36(3): 922-927. Wang Y F, Zhao X L, He B H, et al. Canonical correspondence analysis of summer phytoplankton community and its environmental factors in Hanfeng Lake[J]. Environmental Science, 2015, 36(3): 922-927. |
| [23] | Lv H, Yang J, Liu L M, et al. Temperature and nutrients are significant drivers of seasonal shift in phytoplankton community from a drinking water reservoir, subtropical China[J]. Environmental Science and Pollution Research, 2014, 21(9): 5917-5928. DOI:10.1007/s11356-014-2534-3 |
| [24] | Li R N, Chen Q W, Zhang X Q, et al. Effects of temperature and macronutrients on phytoplankton communities across three largely different lakes identified by a time-space trade-off approach[J]. Ecological Informatics, 2015, 29: 174-181. DOI:10.1016/j.ecoinf.2014.09.004 |
| [25] | Jiang Y J, He W, Liu W X, et al. The seasonal and spatial variations of phytoplankton community and their correlation with environmental factors in a large eutrophic Chinese lake (Lake Chaohu)[J]. Ecological Indicators, 2014, 40: 58-67. DOI:10.1016/j.ecolind.2014.01.006 |
| [26] |
谭啸, 孔繁翔, 曹焕生, 等. 利用流式细胞仪研究温度对两种藻竞争的影响[J]. 湖泊科学, 2006, 18(4): 419-424. Tan X, Kong F X, Cao H S, et al. Influences of temperatures on the competition between two species of algae assayed by flow cytometry[J]. Journal of Lake Sciences, 2006, 18(4): 419-424. DOI:10.18307/2006.0415 |
| [27] | Deng J M, Qin B Q, Paerl H W, et al. Effects of nutrients, temperature and their interactions on spring phytoplankton community succession in Lake Taihu, China[J]. PLoS One, 2014, 9(12): e113960. DOI:10.1371/journal.pone.0113960 |
| [28] | Davis T W, Berry D L, Boyer G L, et al. The effects of temperature and nutrients on the growth and dynamics of toxic and non-toxic strains of Microcystis during cyanobacteria blooms[J]. Harmful Algae, 2009, 8(5): 715-725. DOI:10.1016/j.hal.2009.02.004 |
| [29] |
朱广伟, 金颖薇, 任杰, 等. 太湖流域水库型水源地硅藻水华发生特征及对策分析[J]. 湖泊科学, 2016, 28(1): 9-21. Zhu G W, Jin Y W, Ren J, et al. Characteristics of diatom blooms in a reservoir-water supply area and the countermeasures in Taihu Basin, China[J]. Journal of Lake Sciences, 2016, 28(1): 9-21. DOI:10.18307/2016.0102 |
| [30] | Sugie K, Kuma K, Fujita S, et al. Importance of intracellular Fe pools on growth of marine diatoms by using unialgal cultures and on the Oyashio region phytoplankton community during spring[J]. Journal of Oceanography, 2011, 67(2): 183-196. DOI:10.1007/s10872-011-0017-4 |
| [31] |
姚波, 席北斗, 胡春明, 等. 铁限制对浮游植物生长和群落组成的影响研究综述[J]. 生态环境学, 2010, 19(2): 459-465. Yao B, Xi B D, Hu C M, et al. A review of iron limitation on the growth and competition of phytoplankton[J]. Ecology and Environmental Sciences, 2010, 19(2): 459-465. |