 2017, Vol. 38
2017, Vol. 38



2. 中国地质科学院岩溶地质研究所/国土资源部、广西岩溶动力学重点实验室, 桂林 541004
2. Key Laboratory of Karst Dynamics, Ministry of Land and Resources/Guangxi, Institute of Karst Geology, Chinese Academy of Geological Sciences, Guilin 541004, China
河流作为连接陆地与海洋两大碳库的重要通道, 研究其碳输送是全球碳循环研究的一个重要环节[1].根据组成形态, 河流碳一般可分为溶解无机碳(dissolved inorganic carbon, DIC)、溶解有机碳、颗粒有机碳、颗粒无机碳(particulate inorganic carbon, PIC), 4种形式的碳在水体内部相互转化:PIC可溶解成为DIC, DOC可降解为DIC, 水生植物生长可利用DIC生成POC、DOC[2].每年由陆地生态系统通过河流向海洋排放的有机碳约0.4 Pg, 其中约60%为溶解有机碳(dissolved organic carbon, DOC), 40%是颗粒有机碳(particulate organic carbon, POC)[3], 通过河流陆地生态系统向海洋排放的有机碳大致相当于全球陆地生态系统净初级生产力的1%~2%[4].溶解有机碳根据生物可利用性分为三类:容易被降解的活性DOC(labile DOC, LDOC)、可被缓慢降解的半活性DOC(semi-labile DOC, SLDOC), 以及难以被生物降解的惰性DOC(recalcitrant DOC, RDOC)[5].
岩溶地质作用的碳汇一度被质疑[6], 是源于传统的碳循环模型中把地质作用当作一种纯无机过程[7], 然而, 近来的研究表明, 岩溶作用并非纯无机地质过程, 生物广泛参与岩溶作用, 当水生系统的光合作用强度超过呼吸作用强度时, 碳酸钙沉降过程将无机碳转化成有机碳[8~12], 进一步沉积和埋藏; 同时海洋研究中发现, AAPB(aerobic anoxygenic phototrophic bacteria)作用下形成RDOC, 使碳酸盐岩的风化也能形成长久的碳汇(千年尺度)[13, 14], 通过河流陆地生态系统向海洋排放的有机碳约0.1Pg的DOC循环周期可达到4~6千年[15].
河流有机碳来源主要包括内源有机碳(水生植物光合作用产物)和外源有机碳(地表径流的侵蚀冲刷而进入河流的产物).碳同位素示踪、水中有机质C/N及生物标记法是国内外常用的研究方法, 能很好地解决河流有机碳的来源问题. Waterson等[16]利用C/N和δ13CTOC揭示了美国密西西比河水的总有机碳中50%是内源有机碳, 唐文魁等[2]利用δ13Cpoc研究发现桂江流域水生光合产物对颗粒有机碳的贡献达25.1%, 陶贞等[17]利用C/N研究发现增江流域河水悬移质中的有机碳以水生藻类(内源碳)的贡献为主(超过70%), Sun等[18]利用C/N和δ13CDIC、δ13Cpoc研究发现, 我国西江流域雨季和旱季时河水中3%~21%和12%~22%的POC是河流水生系统的光合作用利用碳酸岩盐风化产物DIC而形成的.原雅琼[19]利用C/N、δ13CPOC分析发现漓江水体在非洪水过程时, 河流有机碳主要来源于水生植物, 内源有机碳的比例可达92%, 且在内源有机碳中来自植物光合利用HCO3-生成的有机碳的比例达46%~77%.原雅琼利用的C/N值与其他数据不是同一时间段, 同时采样密度不够多, DOC与POC中内源有机碳的含量及比例也未做研究, 因此, 剖析水生光合生物对不同形式碳的作用和影响, 有助于对漓江流域碳循环的认识.
本文以珠江支流桂江的上游河段漓江为研究对象, 采用同位素示踪法及相关分析法, 利用稳定同位素、C/N、端元混合模型来辨析漓江流域DOC、POC来源端元及贡献, 分析水生光合生物对流域有机碳的影响, 以期为岩溶碳汇稳定性科学问题的解答提供基础.
1 材料与方法 1.1 研究区概况漓江流域位于广西壮族自治区的东北部, 属珠江水系的桂江上游段.发源于越城岭老山界南侧, 由北向南经兴安、桂林、阳朔, 桂林断面以上河段主要为花岗岩、碎屑岩非岩溶区, 桂林到阳朔河段主要为覆盖型及裸露型岩溶区.地理坐标为E109°45′~111°02′, N24°16′~26°21′, 全长164 km, 流域总面积12 680 km2, 整个漓江流域以漓江为轴线, 呈南北向狭长带状分布(图 1), 属于中亚热带季风气候区, 年平均气温为16.5~20.0℃, 雨量充沛, 年平均降雨量为1 367.5~1 932.9 mm, 雨热同期.漓江为雨源型河流, 河道径流由流域降雨补给, 汛期为每年的4~8月, 其降雨量占全年降雨量的70%左右, 枯季(11月至翌年2月)降雨稀少[20].漓江的地表径流来源于流域内的地表水和地下水, 在雨季发洪水时地表水向地下渗透, 低水期和枯水期地下水补给河槽[20].因流域内碳酸盐岩质纯层厚, 加之雨热同期的季风气候条件, 岩溶发育强烈, 碳酸盐岩峰丛、峰林地貌广布, 漓江贯穿于两者之间, 形成流域区独特的自然景观.

|
图 1 研究区水文地质图与采样点位置 Fig. 1 Hydrogeological map of the study area and the locations of the sampling sites |
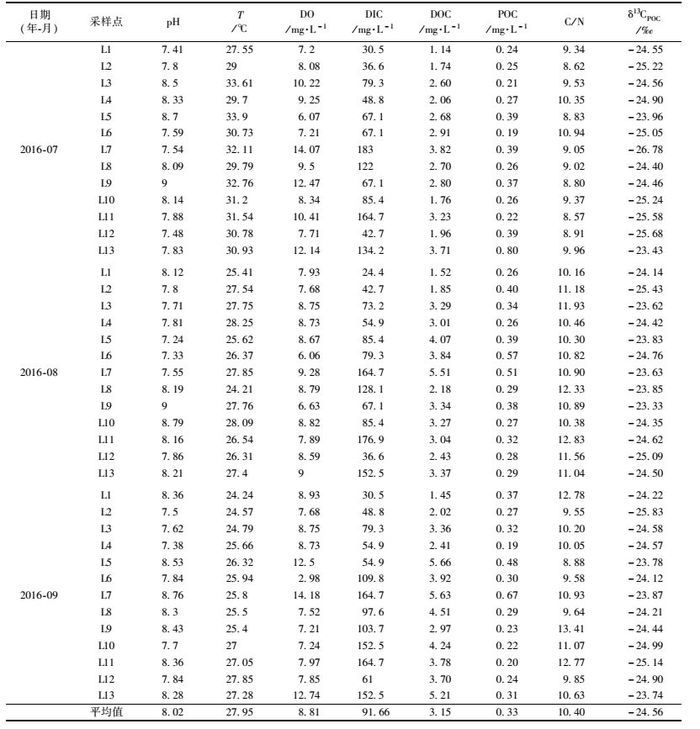
在漓江兴安—阳朔段, 上游(兴安段)、中游(桂林段)和下游(阳朔段)主要断面及支流汇入处布置了13个采样点(图 1), 分别为漓江上游华江(L1)、峡背(L2:灵渠与大溶江汇合后)、灵川(L3:漓江支流甘棠江)、大面圩(L4:甘棠江水汇入漓江后)、漓江干流桂林水文站(L5)、父子岩(L6干流良丰河水汇入漓江后)、漓江支流良丰河(L7)、潮田河水文站(L8:漓江支流潮田河)、省里(L9:干流潮田河水汇入漓江后)、下游干流杨堤(L10)、遇龙河(L11)、遇龙河支流(L12)、阳朔水文站(L13).于2016年7月27日、8月27日、9月12日采集POC、DOC、δ13CPOC、C/N水样, 为避免采样过程中发生污染, 采样前先用待采水样洗涤采样瓶3~5次.现场利用便携式水质分析仪(法国PONSEL)测定水样pH、水温(T)和溶解氧(DO),见表 1, 其精度分别为0.01 pH单位, 0.01℃, 0.01 mg·L-1; 用德国Merck公司生产的碱度计现场滴定HCO3-, 精度为0.1 mmol·L-1.
|
|
表 1 漓江地表水体部分测试数据 Table 1 Part of the testing data of the Lijiang surface water |
1.3 样品分析
DOC含量分析使用德国耶拿公司(Analytik Jena AG)生产的Multi C/N 3100测定, 精度为0.001 mg·L-1. POC和13CPOC的采样及测定:用于过滤水样的玻璃纤维膜预先在马弗炉450℃灼烧6 h以去除无机碳.用玻璃抽滤器加47 mm GF/F滤膜(孔径0.7 μm)过滤, 并送国家海洋局第三海洋研究所测试中心测试, 分析精度为±0.2‰, 测试流程如下:用不锈钢打孔器取固定面积酸熏后的玻璃纤维膜样, 用5×9锡杯包样, 用Thermo公司生产的元素分析仪-稳定同位素质谱仪联机(Flash EA 1112 HT-Delta V Advantages)测定膜样中POC、13CPOC值.载气He流速90 mL·min-1, 反应管温度960℃, 色谱柱温度50℃.
δ13CPOC值以PDB国际标准作为参考标准, δ13CPOC值按以下公式计算:
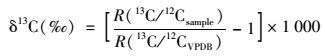
|
(1) |
式中, R(13C/12CVPDB)为国际标准物VPDB(vienna peedee belemnite)的碳同位素丰度比值.
2 结果与讨论 2.1 溶解无机碳与有机碳浓度变化特征本研究测得漓江流域DIC浓度介于24.4~183.0 mg·L-1, 平均为91.66 mg·L-1. DOC浓度介于1.14~5.66 mg·L-1, 平均为3.15 mg·L-1, 低于全球DOC浓度平均值(5.29 mg·L-1)[21]. POC浓度介于0.19~0.8 mg·L-1, 平均为0.33 mg·L-1.
漓江流域DIC浓度空间分布特征为:岩溶区(L7、L11)>岩溶区与非岩溶区的混合区(L3、L4、L5、L6、L9、L8、L10、L13)>非岩溶区(L1、L2、L12), 混合区干流DIC浓度从上游到下游递增, 支流DIC浓度在干流DIC浓度的上下摆动(图 2), 主要受控于流域碳酸盐岩的空间分布比例, DIC质量浓度与碳酸盐岩的空间分布比例呈显著正相关关系(R2=0.80), DIC质量浓度随着碳酸盐岩的分布比例增加而相应增大(图 3). POC含量上下游差异不大(图 2).漓江水体DOC浓度空间分布特征为:岩溶区(3.50 mg·L-1)高于非岩溶区(1.80 mg·L-1); 上游(1.91mg·L-1)低于下游(3.58mg·L-1)(图 2).其原因可能为非岩溶区水进入岩溶区后, 水生植物增加, 水生植物利用HCO3-进行光合作用, 从而导致水体中DOC浓度岩溶区大于非岩溶区. DO也表现为岩溶区大于非岩溶区, 主要是岩溶区水生植物光合作用的原因.一般情况下, 小河流或大河的上游水流湍急, 不利于水生植物的生长; 在大河的下游或河口区, 水流较慢, 营养物质丰富, 有利于河流浮游生物的生长, 因而下游较上游有较高的水生生物初级生产力[22].

|
图 2 漓江流域DIC、DOC、POC浓度变化特征 Fig. 2 Variations in content of DIC, DOC, and POC in the Lijiang basin |

|
图 3 DIC与碳酸盐岩分布比例相关关系 Fig. 3 Correlation between DIC and distribution proportion of carbonate rocks |
漓江流域DOC浓度与DIC质量浓度存在正相关关系(图 4), 说明DOC来源除了外源, 还存在DIC向DOC的转化; DOC浓度与DO存在正相关关系(图 4), 说明DOC和DO有共同的来源(水生生物光合作用).综上, 说明漓江岩溶地表水生系统中水生植物的光合作用可以利用DIC产生DOC, 其过程表示如下:

|
图 4 DOC与DIC及DOC与DO的相关关系 Fig. 4 Correlation between DOC and DIC, DOC, and DO |

|
(2) |
漓江水体POC浓度与DO呈正相关关系(图 5), 说明POC来源除了外源, 还可能来源于水生生物的初级生产力; POC浓度与DOC存在正相关关系(图 5), 说明漓江岩溶地表水生系统可能存在DOC与POC之间的转化.

|
图 5 POC与DO、POC与DOC的相关关系 Fig. 5 Correlation between POC and DO, POC, and DOC |
漓江流域13CPOC值介于-26.78‰~-23.33‰, 平均为-24.56‰, 上下游差异不明显.河流输送的颗粒有机碳可以看作水生生物的生物量(内源碳)与外源土壤颗粒有机碳的端元混合.考虑内源、外源端元组分的13CPOC, 可以定量估算河水POC中内源碳与外源碳的比例, 其同位素质量平衡方程如下:
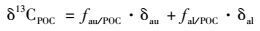
|
(3) |

|
(4) |
式中, 13CPOC为样品的碳同位素值; δau、δal分别为内源碳与外源碳端元组分的碳同位素值; fau/POC、fal/POC分别表示POC中内源碳与外源碳所占的比例.
两个端元的确定如下:其中δal可以参照Sun等[18]关于西江2005年13CPOC的数据, 6月西江发生罕见洪水, 期间13CPOC介于-23.8‰~-22.2‰, 平均为-23.0‰, 河流初级生产对POC的贡献可以忽略不计, POC几乎全部来自于陆源侵蚀, 此时的13CPOC可以作为δal的典型值, 为-23.0‰; δau则选择Sun在西江下游水生生物量的13C取值, 为-32‰.
结合POC浓度值, 可计算出漓江流域颗粒有机碳中内源有机碳浓度, 方程如下:
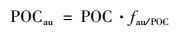
|
(5) |
式中, POC为样品的POC浓度值; fau/POC为POC中内源碳所占比例; POCau为POC中内源有机碳的浓度.
计算结果表明水生生物量对漓江流域POC贡献率范围为3.69%~41.94%, 平均为17.31%, 低于桂江水体水生生物量对POC的贡献7月平均值(25.1%)[2]; POC中内源有机碳浓度(POCau)介于0.01~0.16 mg·L-1, 平均为0.05mg·L-1(图 6、图 7).总体而言, 漓江流域POC以外源有机碳为主, 上下游差异不大.即漓江水体中83%的POC来源于陆生生物, 由水生生物转化过来的POC占的比例为17%.

|
图 6 漓江流域TOC、DOC、POC中内源碳的含量 Fig. 6 Content of autochthonous organic carbon in the TOC, DOC, and POC of the Lijiang basin |

|
图 7 漓江流域TOC、DOC、POC中内源碳的比例 Fig. 7 Proportion of allochthonous organic carbon in the TOC, DOC, and POC of the Lijiang basin |
本研究测得漓江流域C/N值介于8.57~13.41之间, 平均为10.40.已有研究表明外源有机碳C/N值大于15[23]; 根据典型藻类光合作用形成产物为C5.7H9.8O2.3N, 其C/N应为6左右, 真菌(C10H17O6N)的C/N为10, 细菌(C5H7O2N)的C/N为5, 因此, 内源有机质的C/N介于5~10之间.以C/N值6.6为内源端元[24, 25], 20.69为外源端元[26], 可以利用端元混合模型计算内源有机碳占总有机碳的比例, 方程如下:
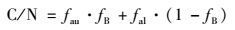
|
(6) |
式中C/N为样品的C/N值; fau为C/N值的内源端元, 取6.6; fal为C/N值的外源端元, 取20.69; fB为内源有机碳在总有机碳中的比例.
结合漓江流域TOC(POC+DOC)浓度值, 可计算出总有机碳中内源碳的浓度(TOCau), 方程如下:
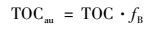
|
(7) |
式中, TOC为样品的总有机碳含量值; fB为内源有机碳在总有机碳中的比例; TOCau为总有机碳中内源碳的浓度.
计算表明内源有机碳占总有机碳的比例(fB)介于51.68%~85.99%, 平均为73.07%;总有机碳中内源碳的含量介于0.97~5.10mg·L-1, 平均为2.48mg·L-1(图 6、图 7).即漓江水体中27%的TOC来源于陆源, 73%的TOC来源于水生植物光合作用产物.
2.4 DOC、TOC、POC内外源比例及含量分析结合总有机碳中内源碳的浓度、颗粒有机碳中内源碳的浓度、溶解有机碳浓度, 可以计算出DOC中内源碳的浓度(DOCau)及DOC中内源碳所占比例(fau/DOC).计算公式如下:

|
(8) |

|
(9) |
式中, DOCau为DOC中内源碳的浓度; TOCau为总有机碳中内源碳的浓度; POCau为POC中内源碳的浓度; DOC为样品的溶解有机碳浓度; fau/DOC为DOC中内源碳所占比例.
计算结果表明漓江水体DOC中内源碳所占比例(fau/DOC)介于54.43%~94.69%, 平均为79.51%; DOC中内源碳的浓度介于0.97~5.10 mg·L-1, 平均为2.48 mg·L-1.即漓江水体中20%的DOC是地表径流的侵蚀冲刷而进入河流的产物, 80%的DOC来源于水生生物的初级生产力.
漓江水体内源DOC占内源TOC的比例介于92.83%~99.46%, 平均为97.58%, 空间分布无明显差异; DOC占TOC的比例平均为89.83%(DOC是构成有机碳的主体), 空间分布表现为岩溶区、混合区>非岩溶区(图 8). DOC浓度空间分布特征为:岩溶区>混合区>非岩溶区, POC浓度空间分布差异不大(图 2).内源TOC、DOC浓度空间分布特征为:岩溶区>混合区>非岩溶区, 内源POC浓度空间分布差异不大(图 6), 可能与岩溶区水生植物丰茂、碳酸酐酶(carbonic anhydrase, CA)活性较强有关(CA活性与HCO3-浓度呈正相关)[27]; 漓江水体TOC、DOC、POC中内源碳的比例空间分布差异不大(图 7); TOC来源以内源碳为主(内源碳的比例为73%), 而POC主要来源于陆生生物(内源碳的比例为17%), DOC主要来源于水生生物的初级生产力(内源碳的比例为80%).

|
图 8 DOC、POC占TOC比例 Fig. 8 Proportion of DOC and POC in the TOC |
综上所述漓江流域TOC、DOC以内源碳为主, POC中水生生物量也占一定比例.
内源有机碳中有多少比例是水生生物利用HCO3-生成的、内源有机碳的稳定性及DOC与POC之间的转化本文都未做研究, 以后可以展开这方面的研究.
3 结论(1) 漓江流域DIC浓度空间分布特征为:岩溶区(良丰河、遇龙河)>岩溶区与非岩溶区的混合区(灵川、大面圩、桂林水文站、父子岩、潮田河水文站、省里、杨堤、阳朔水文站)>非岩溶区(华江、峡背、遇龙河支流); 混合区干流DIC含量从上游到下游递增.主要受控于流域碳酸盐岩的空间分布比例, DIC质量浓度与碳酸盐岩的空间分布比例呈显著正相关关系(R2=0.80, P < 0.01).
(2) 漓江水体DOC浓度与DIC质量浓度、DOC浓度与DO均存在正相关关系, 说明岩溶地表水生系统中水生植物的光合作用可以利用DIC而形成DOC; POC浓度与DO呈正相关关系, 说明POC来源除了外源, 还可能来源于水生生物的初级生产力; POC浓度与DOC存在正相关关系, 岩溶地表水生系统存在DOC与POC转化.
(3) POC浓度、POC中内源碳的浓度及POC中内源碳的比例空间分布差异不大, POC来源以外源碳为主, POC中内源有机碳浓度介于0.01~0.16mg·L-1, 平均为0.05mg·L-1, 水生生物量对漓江流域POC贡献介于3.69%~41.94%, 平均为17.31%, 即83%的POC来源于陆生生物, 由水生生物转化过来的POC占的比例为17%.
(4) DOC是构成漓江水体TOC的主体, TOC来源以内源有机碳为主, 内源碳的浓度空间分布特征为:岩溶区>混合区>非岩溶区, 可能与岩溶区水生植物丰茂、碳酸酐酶活性较强有关, TOC中内源碳的浓度介于1.02~5.14 mg·L-1, 平均为2.54 mg·L-1; TOC中内源碳的比例空间分布差异不大, 介于51.68%~85.99%, 平均为73.07%, 即27%的TOC来源于陆源, 73%的TOC来源于水生植物光合作用产物.
(5) DOC浓度、内源DOC浓度空间分布均为岩溶区>混合区>非岩溶区, DOC主要来源于水生生物的初级生产力, DOC中内源碳的浓度介于0.97~5.10 mg·L-1, 平均为2.48 mg·L-1; DOC中内源碳的比例空间分布差异不大, DOC中内源碳所占比例介于54.43%~94.69%, 平均为79.51%, 20%的DOC是地表径流的侵蚀冲刷而进入河流的产物, 80%的DOC来源于水生生物的初级生产力.
| [1] | Alvarez-Cobelas M, Angeler D G, Sánchez-Carrillo S, et al. A worldwide view of organic carbon export from catchments[J]. Biogeochemistry, 2012, 107(1-3): 275-293. DOI:10.1007/s10533-010-9553-z |
| [2] |
唐文魁, 陶贞, 高全洲, 等. 桂江主要离子及溶解无机碳的生物地球化学过程[J]. 环境科学, 2014, 35(6): 2099-2107. Tang W K, Tao Z, Gao Q Z, et al. Biogeochemical processes of the major ions and dissolved inorganic carbon in the Guijiang river[J]. Environmental Science, 2014, 35(6): 2099-2107. |
| [3] | Spitzy A, Ittekkot V. Dissolved and particulate organic matter in rivers[A]. In: Mantoura R F C, Martin J M, Wollast R (Eds.). Ocean Margin Processes in Global Change[M]. Chichester: John Wiley, 1991. 5-17. |
| [4] | Meybeck M. Carbon, nitrogen, and phosphorus transport by world rivers[J]. American Journal of Science, 1982, 282(4): 401-450. DOI:10.2475/ajs.282.4.401 |
| [5] |
焦念志. 海洋固碳与储碳——并论微型生物在其中的重要作用[J]. 中国科学:地球科学, 2012, 42(10): 1473-1486. Jiao N Z. Carbon fixation and sequestration in the ocean, with special reference to the microbial carbon pump[J]. Scientia Sinica Terrae, 2012, 42(10): 1473-1486. |
| [6] | Curl R L. Carbon shifted but not sequestered[J]. Science, 2012, 335(6069): 655. |
| [7] | Berner R A, Lasaga A C, Garrels R M. The carbonate-silicate geochemical cycle and its effect on atmospheric carbon dioxide over the past 100 million years[J]. American Journal of Science, 1983, 283(7): 641-683. DOI:10.2475/ajs.283.7.641 |
| [8] | Zhang C, Wang J L, Pu J B, et al. Bicarbonate daily variations in a karst river: the carbon sink effect of subaquatic vegetation photosynthesis[J]. Acta Geologica Sinica, 2012, 86(4): 973-979. DOI:10.1111/j.1755-6724.2012.00721.x |
| [9] | Jiang Y J, Hu Y J, Schirmer M. Biogeochemical controls on daily cycling of hydrochemistry and δ13 C of dissolved inorganic carbon in a karst spring-fed pool[J]. Journal of Hydrology, 2013, 478: 157-168. DOI:10.1016/j.jhydrol.2012.12.001 |
| [10] | Tobias C, Böhlke J K. Biological and geochemical controls on diel dissolved inorganic carbon cycling in a low-order agricultural stream: implications for reach scales and beyond[J]. Chemical Geology, 2011, 283(1-2): 18-30. |
| [11] | de Montety V, Martin J B, Cohen M J, et al. Influence of diel biogeochemical cycles on carbonate equilibrium in a karst river[J]. Chemical Geology, 2011, 283(1-2): 31-43. |
| [12] |
刘再华, 李强, 孙海龙, 等. 云南白水台钙华水池中水化学日变化及其生物控制的发现[J]. 水文地质工程地质, 2005, 32(6): 10-15. Liu Z H, Li Q, Sun H L, et al. Diurnal variations in hydrochemistry in a travertine-depositing stream at Baishuitai, Yunnan, SW China: observations and explanations[J]. Hydrogeology and Engineering Geology, 2005, 32(6): 10-15. |
| [13] | Liu Z H, Dreybrodt W, Wang H J. A new direction in effective accounting for the atmospheric CO2 budget: considering the combined action of carbonate dissolution, the global water cycle and photosynthetic uptake of DIC by aquatic organisms[J]. Earth-Science Reviews, 2010, 99(3-4): 162-172. DOI:10.1016/j.earscirev.2010.03.001 |
| [14] |
焦念志, 张传伦, 李超, 等. 海洋微型生物碳泵储碳机制及气候效应[J]. 中国科学:地球科学, 2013, 43(1): 1-18. Jiao N Z, Zhang C L, Li C, et al. Controlling mechanisms and climate effects of microbial carbon pump in the ocean[J]. Scientia Sinica Terrae, 2013, 43(1): 1-18. |
| [15] | Bauer J E, Williams P M, Druffel E R M. 14C activity of dissolved organic carbon fractions in the north-central Pacific and Sargasso Sea[J]. Nature, 1992, 357(6380): 667-670. DOI:10.1038/357667a0 |
| [16] | Waterson E J, Canuel E A. Sources of sedimentary organic matter in the Mississippi River and adjacent Gulf of Mexico as revealed by lipid biomarker and δ13 CTOC analyses[J]. Organic Geochemistry, 2008, 39(4): 422-439. DOI:10.1016/j.orggeochem.2008.01.011 |
| [17] |
陶贞, 高全洲, 姚冠荣, 等. 增江流域河流颗粒有机碳的来源、含量变化及输出通量[J]. 环境科学学报, 2004, 24(5): 789-795. Tao Z, Gao Q Z, Yao G R, et al. The sources, seasonal variation and transported fluxes of the riverine particulate organic carbon of the Zengjiang River, Southern China[J]. Acta Scientiae Circumstantiae, 2004, 24(5): 789-795. |
| [18] | Sun H G, Han J T, Zhang S R, et al. Transformation of dissolved inorganic carbon (DIC) into particulate organic carbon (POC) in the lower Xijiang River, SE China: an isotopic approach[J]. Biogeosciences Discussions, 2011, 8(5): 9471-9501. DOI:10.5194/bgd-8-9471-2011 |
| [19] |
原雅琼. 水生光合生物对漓江流域水化学和岩溶碳汇的影响[D]. 重庆: 西南大学, 2016. 1-69. Yuan Y Q. Impacts of aquatic organism on hydrochemical characteristics and karst carbon sink in Lijiang basin[D]. Chongqing: Southwest University, 2016. 1-69. |
| [20] | 韩耀全, 周解, 吴祥庆. 漓江的自然地理与水质调查[J]. 广西水产科技, 2007(2): 8-16. |
| [21] | Dai M H, Yin Z Q, Meng F F, et al. Spatial distribution of riverine DOC inputs to the ocean: an updated global synthesis[J]. Current Opinion in Environmental Sustainability, 2012, 4(2): 170-178. DOI:10.1016/j.cosust.2012.03.003 |
| [22] |
张永领. 河流有机碳循环研究综述[J]. 河南理工大学学报(自然科学版), 2012, 31(3): 344-351. Zhang Y L. The review of the research of the riverine organic carbon cycle[J]. Journal of Henan Polytechnic University (Natural Science), 2012, 31(3): 344-351. |
| [23] | Kendall C, Silva S R, Kelly V J. Carbon and nitrogen isotopic compositions of particulate organic matter in four large river systems across the United States[J]. Hydrological Processes, 2001, 15(7): 1301-1346. DOI:10.1002/(ISSN)1099-1085 |
| [24] | Redfield A C. The biological control of chemical factors in the environment[J]. American Scientist, 1958, 46(3): 230A-205-221. |
| [25] | LaZerte B D. Stable carbon isotope ratios: implications for the source of sediment carbon and for phytoplankton carbon assimilation in Lake Memphremagog Quebec[J]. Canadian Journal of Fisheries and Aquatic Sciences, 1983, 40(10): 1658-1666. DOI:10.1139/f83-192 |
| [26] |
邢长平. 华南亚热带森林土壤14C年龄及14C示踪研究[D]. 广州: 中国科学院广州地球化学研究所, 1998. 1-57. Xing C P. Studies on 14C age and 14C tracing for subtropical forest soils in South China[D]. Guangzhou: Guangzhou Institute of Geochemistry, Chinese Academy of Science, 1998. 1-57. |
| [27] |
申泰铭, 李为, 张强, 等. 流域不同地质生态环境中水体碳酸酐酶活性特征——以桂江流域为例[J]. 中国岩溶, 2012, 31(4): 409-414. Shen T M, Li W, Zhang Q, et al. Carbonic anhydrase activity of the water-body in different eco-environments of river basins: a case study in the Guijiang river basin[J]. Carsologica Sinica, 2012, 31(4): 409-414. |