 2017, Vol. 38
2017, Vol. 38


2. 西北水资源与环境生态教育部重点实验室, 西安 710055;
3. 浙江工业大学环境学院, 杭州 310014
2. Key Laboratory of Northwest Water Resource, Environment and Ecology, Ministry of Education, Xi'an 710055, China;
3. College of Environment, Zhejiang University of Technology, Hangzhou 310014, China
厌氧氨氧化(anaerobic ammonium oxidation, ANAMMOX)是指在厌氧或缺氧条件下,厌氧氨氧化菌以亚硝氮为电子受体将氨氮氧化为氮气的过程[1~3].与传统硝化/反硝化过程相比,厌氧氨氧化反应可减少了100%的曝气能耗、100%的有机碳源、80%~90%的污泥产率[4, 5],因此厌氧氨氧化反应被称为一种高效低耗的生物脱氮技术.然而厌氧氨氧化菌生长缓慢(μmax=0.065 d-1)世代时间长(11 d)[6]成为制约厌氧氨氧化工艺广泛应用的主要原因,因此如何快速启动厌氧氨氧化工艺成为突破其应用的一种有效手段.其次厌氧氨氧化菌对环境变化十分敏感,其活性容易受到抑制且恢复周期长,因此找到可反映厌氧氨氧化反应性能的简单指标对维持反应器稳定运行十分必要.研究发现厌氧氨氧化颗粒污泥不仅可以提高污泥的沉降性还有利于提高厌氧氨氧化菌对环境的适应性,认为培养厌氧氨氧化颗粒污泥是维持厌氧氨氧化菌浓度的手段之一[7, 8].
已有很多研究表明胞外聚合物(extracellular polymeric substances, EPS)在污泥颗粒化及维持颗粒污泥的结构稳定中起到重要作用[9, 10].例如EPS中的蛋白质(PN)和多糖(PS)直接影响细胞表面电性、亲疏水性及生物聚集体的空间结构[11, 12].虽然研究发现厌氧氨氧化菌可分泌较多的EPS[13],但目前国内外关于EPS的研究主要集中在好氧颗粒污泥领域[14, 15],针对厌氧氨氧化颗粒污泥的EPS研究较少.现有关于厌氧氨氧化菌EPS的研究主要集中在EPS的组成、含量[16, 17],关于EPS在厌氧氨氧化颗粒污泥内的空间分布及其在厌氧氨氧化污泥颗粒化过程中的作用的研究较少.因此有必要研究EPS在厌氧氨氧化污泥颗粒化过程中的作用及其在厌氧氨氧化菌细胞外的分布.
本研究采用厌氧序批式反应器(anaerobic sequencing batch reactor, ASBR)成功富集厌氧氨氧化颗粒污泥,解释了以活性污泥做接种泥时厌氧氨氧化反应器的启动过程,探讨了表征厌氧氨氧化反应器性能的简单指标.采用多重荧光染色的方法考察了EPS在厌氧氨氧化颗粒污泥内的空间分布,进一步解释了EPS的不同组分在厌氧氨氧化污泥颗粒化中作用,以期为快速判断厌氧氨氧化反应器的运行情况提供理论依据,并直观地解释了EPS对形成厌氧氨氧化颗粒污泥的作用.
1 材料与方法 1.1 实验装置与运行本实验采用ASBR反应器,反应器工作容积为5 L.反应器通过外部加热水槽及温控系统装置控制反应温度为35℃±1℃.反应器的进水、反应、沉淀和出水由PLC控制自动运行.启动初期,水力停留时间(hydraulic retention time, HRT)为36 h,反应器稳定运行后,HRT缩短至18 h.稳定运行周期为8 h,其中进水360 min;反应90 min;沉淀15 min;出水10 min;闲置5 min.搅拌速度为70 r·min-1,出水pH控制7.5~8.0.
1.2 实验用水及接种污泥反应器运行期间采用人工配水,进水组分见表 1,微量元素溶液[18]组分见表 2.根据出水pH值采用1 mol·L-1 HCl和1 mol·L-1 NaOH调节进水pH.接种污泥取自实验室稳定运行的A/A/O系统的活性污泥.
|
|
表 1 实验废水组分 Table 1 Wastewater composition |
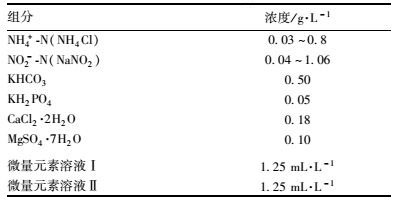
|
|
表 2 微量元素组分 Table 2 Microelement composition |
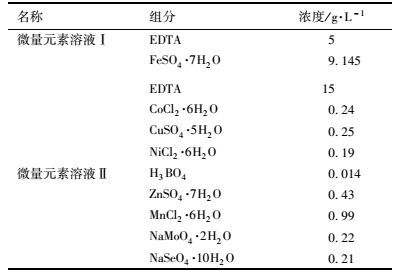
1.3 分析方法 1.3.1 常规指标分析
NH4+-N采用纳氏试剂分光光度法;NO2--N采用N-(1-萘基)-乙二胺分光光度法;NO3--N采用紫外分光光度法;pH:雷磁PHS-3C型pH计;MLSS和MLVSS采用重量法.此外,厌氧氨氧化反应器产气量采用湿式气体流量计进行计量.
1.3.2 荧光原位杂交(fluorescent in situ hybridization, FISH)样品预处理:取反应器活性污泥2 mL,离心(10 000 r·min-1)5 min,去上清液,加入1×PBS缓冲溶液1 mL重悬,重复操作两次后,加入1 mL的4%多聚甲醛溶液重悬,放置4℃下固定3 h,使细胞形态固定.然后离心,倾去上清液,加入1×PBS缓冲溶液1 mL离心5 min,洗去多余的多聚甲醛溶液重复操作3次.最后,分别加入体积比为1:1的1×PBS缓冲溶液和无水乙醇,摇匀置于-20℃保存.
脱水和杂交:取由4%多聚甲醛固定后的污泥样品,均匀涂抹于经酒精清洗过的载玻片上,至少取3个平行样,干燥后的样品依次用50%、80%、95%的乙醇溶液各浸泡3 min对细胞进行脱水,取出后风干.配2 mL杂交缓冲液(SDS 10%,Tris-HCl 1 mol·L-1,NaCl 5 mol·L-1,去离子甲酰胺,ddH2O).将无菌水配置成的25 ng·μL-1的探针与杂交缓冲液按体积比1:8混合成探针使用液,用锡箔纸包好放入46℃中预热5 min.将剩余的杂交缓冲液遍布在杂交管内折好的吸水纸上,目的是让样品在杂交过程中始终处于杂交缓冲液的潮湿环境中,将固定好样品的载玻片放入杂交管中,在46℃中放置数分钟.取9 μL均匀滴入载玻片上的污泥试样中,然后将载玻片迅速移回杂交管中在46℃中避光杂交2~3 h.杂交结束后,将载玻片快速放入预热后的淋洗缓冲液(SDS 10%,Tris-HCl 1 mol·L-1,NaCl 5 mol·L-1,ddH2O)中,46℃水浴10 min后用4℃冰水冲洗样品,于黑暗中自然风干.经过风干的样品封片后使用激光共聚焦显微镜(Leica TCS SP8) 进行观察.
本实验所用探针(表 3):总细菌、厌氧氨氧化菌分别采用Eub338mix、Amx368.
|
|
表 3 荧光原位杂交实验中所用的探针 Table 3 Probes used in FISH |

1.3.3 胞外聚合物的提取及测定
EPS的提取采用阳离子交换树脂法(cation exchange resin, CER)[22].取含有0.1 g MLVSS的污泥样品,4℃、5 000 r·min-1转速下离心10 min后弃去上清液.将上述离心产生的泥饼用PBS缓冲液(含有2 mmol·L-1Na3PO4,4 mmol·L-1NaH2PO4,9 mmol·L-1 NaCl和1 mmol·L-1KCl,并调节pH为7) 重悬至30 mL.然后将重悬后的混合液在4℃、5 000 r·min-1转速下离心10 min.此过程重复两次,以确保获得干净的污泥.称取7 g阳离子交换树脂(Dower Marathon C,Na+型,20~50目,货号Fluka 91973) 于锥形瓶中,加入30 mL PBS缓冲液,于磁力搅拌器800 r·min-1搅拌1 h,弃去上清液(清洗树脂过程).随后将上述洗净的污泥用分散机分散5 min,并用PBS缓冲液冲洗至30 mL,转移到已洗好的树脂中,闭光800 r·min-1搅拌4 h进行提取.提取过程结束后,将混合液4℃、5 000 r·min-1离心10 min;将离心产生的上清液再次离心15 min,4℃、12 000 r·min-1.
蛋白质的测定采用Lowry法,以牛血清蛋白作为标准物质,具体步骤参照Frølund等[22]的描述进行;多糖的测定采用苯酚-硫酸法,以无水葡萄糖作为标准物质,具体步骤参照DuBois等[23]的描述进行.
1.3.4 多重荧光染色分析为了对厌氧氨氧化颗粒污泥中的蛋白质、多糖、脂类及死/活细胞的相对含量和空间分布进行分析,本实验使用不同的荧光染料分别对EPS和死/活细胞进行多重荧光染色.异硫氰酸荧光素(FITC)用于胞外蛋白质的染色;Con A(Tetramethylrhodamine conjugates of Concanvalin A)用来指示α-呋喃葡萄糖和α-甘露糖残基;卡尔科弗卢尔荧光增白剂(Calcofluor white)用来染色β-D-呋喃葡萄糖基;尼罗红(Nile red)用以指示脂类及疏水基团;使用SYTO 63染色颗粒污泥中总细胞;SYTOX blue用于死细胞的染色.多重荧光染色所用荧光染料的激发波长、发射波长、用量和使用浓度见表 4.其中FITC和尼罗红购自美国Mpbio公司;Calcofluor white购自美国Sigma公司;Con A、SYTO 63和SYTOX blue购自美国invitrogen公司.
|
|
表 4 颗粒污泥多重荧光染色所用染料特性 Table 4 Stains used in the proposed staining scheme |
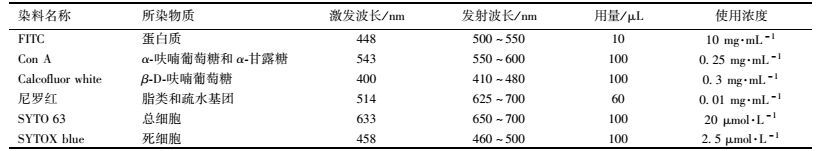
根据Chen等[24]提出的方法,对厌氧氨氧化颗粒污泥进行多重荧光染色.步骤如下:将适量厌氧氨氧化颗粒污泥用pH为7.2,浓度为0.1 mol·L-1的PBS缓冲溶液清洗3遍,首先把100μL的SYTO 63工作液滴加到样品中染色30 min,然后在样品中添加100 μL碳酸氢钠缓冲液(0.1 mol·L-1),使胺基保持非质子化的状态,再将样品用10 μL的异硫氰酸荧光素(FITC)溶液染色1 h,后加入100 μL的ConA溶液染色30 min,再在样品中添加100 μL的Calcofluor white溶液染色30 min,最后加入60 μL尼罗红染色10 min,每次染色后均要用pH=7.2的PBS缓冲溶液清洗3遍.染色完毕后,将样品-4℃保存.在观察前在样品中添加100 μL的SYTOX blue溶液染色5 min.厌氧氨氧化颗粒污泥经染色后,使用冰冻切片机(Leica CM1950) 进行切片(厚度为10~20μm),再使用激光扫描共聚焦显微镜(Leica TCS SP8) 进行观察.
2 结果与讨论 2.1 反应器的启动及运行特性反应器启动后,通过逐步提高进水NH4+-N、NO2--N浓度及缩短HRT的方式提高反应器的氮负荷. 图 1为启动初期至稳定运行期反应器运行效果图.根据出水NH4+-N浓度变化可将整个启动过程分为活性停滞期(0~23 d)、活性提高期(24~122 d)和稳定运行期(123~160 d)这3个阶段.

|
(a)进、出水NH4+-N、NO2--N、NO3--N浓度变化,HRT变化;(b) NH4+-N、NO2--N、TN去除率,进水TN负荷及去除TN负荷 图 1 厌氧氨氧化反应器运行效果 Fig. 1 Performance of the ANAMMOX reactor |
在阶段Ⅰ(0~23 d),出水NH4+-N浓度高于进水NH4+-N浓度,而出水NO2--N浓度低于进水NO2--N浓度,出水NO3--N浓度基本为零.路青等[25]分别接种好氧污泥、厌氧颗粒污泥和厌氧消化污泥均成功启动厌氧氨氧化,在启动初期,出水NH4+-N浓度大于进水,这与本研究结果一致.这是由于污泥接种后微生物所处环境改变,使得接种硝化污泥中的好氧菌因不能适应厌氧环境而发生细胞自溶[26],其次是由于硝化污泥中厌氧氨氧化菌所占份额很小,消耗NH4+-N有限,从而释放出大量的NH4+-N,导致出水NH4+-N浓度升高.此外,细菌自溶产生大量的有机物为反硝化菌提供了电子供体和碳源,反硝化成为该阶段的主反应,进水NO2--N因反硝化作用而得到去除[26].
在阶段Ⅱ(24~120 d),出水NH4+-N浓度逐渐降低,在24~66 d时出水NH4+-N浓度接近进水NH4+-N浓度,随后出水NH4+-N浓度大幅下降.而出水NO2--N浓度先升高后下降,这是由于前一阶段中细菌自溶产生的有机物被反硝化菌利用和进水稀释后,使得反硝化菌失去电子供体而活性减弱,导致NO2--N去除率降低,出水NO2--N浓度升高[27],随着厌氧氨氧化活性的提高,NO2--N去除率升高.在67 d时,出水NH4+-N、NO2--N浓度同时小于进水NH4+-N、NO2--N浓度,首次出现明显的厌氧氨氧化反应.随后厌氧氨氧化菌活性逐渐增大,进水浓度快速提升(67~122 d),此时厌氧氨氧化菌成为优势菌种.
在阶段Ⅲ(123~170 d),反应器厌氧氨氧化菌活性稳定,标志着厌氧氨氧化反应系统启动成功.通过缩短HRT,进一步提高反应器的氮负荷,反应负荷最终稳定在1.92 kg·(m3·d)-1,总氮(TN)去除负荷达到1.7 kg·(m3·d)-1.本阶段内,NH4+-N、NO2--N去除率均接近100%,TN去除率为89.87%±0.43%.
2.2 厌氧氨氧化反应器的性能指标厌氧氨氧化菌生长缓慢、对外界环境变化极其敏感[28].因此,厌氧氨氧化菌的活性容易受到抑制,并且恢复周期长.研究者有必要借助一些简单、高效的性能指标快速判断反应器的运行状况.
2.2.1 pH很多研究者指出厌氧氨氧化反应器出水pH高于进水pH[29~31].如图 2(a)所示,实验过程中发现厌氧氨氧化反应器出水pH远高于进水pH,并且总氮去除负荷与进出水pH差值、进水量(ΔpH×Q)之间存在明显的线性相关性(r2=0.936).出水pH值升高是由于厌氧氨氧化反应过程消耗质子[29].因此如果进水水质和反应器运行参数不变,可借助出水pH值快速判断厌氧氨氧化反应器的运行状况.如果在进水水质恒定的条件下出水pH有很大降低,这就说明反应器运行中出现问题.

|
(a)进、出水pH及ΔpH的变化;(b)去除负荷与ΔpH×Q的关系 图 2 厌氧氨氧化反应器的性能指标(pH) Fig. 2 The pH indicator for start-up performance of the ANAMMOX reactor |
由于厌氧氨氧化菌通过硝酸盐还原酶将NO2--N氧化为NO3--N获得生长所需的能量,因此NO3--N产率也是厌氧氨氧化反应性能的一个重要指标.实验期间发现出水NO3--N浓度与进水TN浓度有良好的线性相关性(图 3).因此如果在进水浓度不变的条件下,出水NO3--N浓度出现很大程度的下降,说明反应器运行出现问题,需要及时处理.

|
图 3 进水TN浓度与出水NO3--N浓度的关系 Fig. 3 Relationship between influent TN concentration and effluent NO3--N concentration |
目前,研究者普遍接受的厌氧氨氧化反应化学计量关系如式(1) 所示[32].然而,不同研究者得出的结论之间存在较大差异,冯平等[33]的研究结果表明,稳定运行阶段三者之间的比值为1:1.44:0.26,而张海芹等[34]采用不同接种污泥启动ABR厌氧氨氧化反应器时发现稳定运行后三者的比值分别为1:1.16:0.21和1:1.27:0.36.除此之外一些研究者发现由于进水基质、运行条件、反应器类型和厌氧氨氧化菌菌种的不同,NO3--N/NH4+-N在0.5~4之间变化[35, 36].本实验中,NH4+-N与NO2--N的消耗量和NO3--N生成量之间的比例关系为1:1.32±0.08:0.24±0.03. NH4+-N与NO2--N消耗量的比值与Strous等[32]提出的理论值(1.32) 一致,而NO3--N生成量与NH4+-N消耗量的比值小于0.26,这可能是由于系统中还存在少量反硝化菌,其利用菌体由于生长过程中死亡或外层组织脱落产生的少量有机物,进行反硝化作用去除一部分NO3--N.
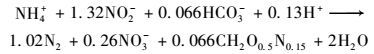
|
(1) |
实验期间采用FISH技术对絮体污泥及颗粒污泥中厌氧氨氧化菌的百分比进行检测.图 4显示的绿色荧光信号(Amx338) 与红色荧光信号(EUB338mix)几乎完全重合,说明此时厌氧氨氧化菌成为反应器中的优势菌,厌氧氨氧化菌占总细菌的87.39%±4.76%.絮体污泥的FISH图片显示厌氧氨氧化菌的荧光信号呈环形,其内部荧光强度较弱,这一结果与Strous等[32]和Araujo等[37]的研究结果一致.这是由于厌氧氨氧化菌细胞内部存在一个被称为厌氧氨氧化体的隔室,而厌氧氨氧化体膜的主要成分为脂类,厌氧氨氧化体内不含核糖体[38].

|
(a)红色EUB338mix; (b)绿色Amx338;(c)a, b单个图像的叠加 图 4 厌氧氨氧化絮体污泥的FISH(bar=10 μm) Fig. 4 FISH images of ANAMMOXA floc sludge(bar=10 μm) |
由颗粒污泥的FISH图片(图 5)可看出,厌氧氨氧化菌主要集中在颗粒污泥外侧,颗粒污泥内部较少.颗粒污泥中厌氧氨氧化菌占总细菌的90.93%±6.89%.这是由于基质受传质阻力影响,颗粒污泥内部基质浓度低,外部基质充足.由图 4和5可看出厌氧氨氧化菌容易形成生物聚集体,这可能是由于厌氧氨氧化菌可分泌大量EPS.

|
(a)红色EUB338mix;(b)绿色Amx338;(c)a, b单个图像的叠加;(d)局部放大图;(a)~(c)bar=40 μm,(d)bar=10 μm 图 5 厌氧氨氧化颗粒污泥的FISH Fig. 5 FISH images of ANAMMOXA granular sludge |
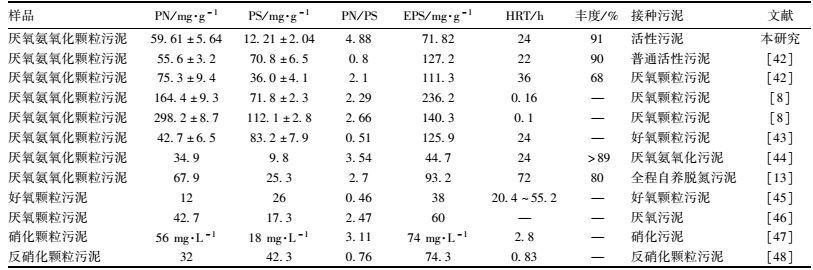
EPS的化学成分比较复杂,其组分因来源不同而有所差别.总体而言,PN和PS是EPS的主要成分[39]. CER法提取后,测得其中蛋白质和多糖的含量分别为(59.61±5.64) mg·g-1和(12.21±2.04) mg·g-1,PN/PS为4.88±1.39.其他关于厌氧氨氧化颗粒污泥EPS的研究也发现PN是EPS的主要组分[7, 8].这可能是由于EPS是细胞体与水溶液间的离子传递通道,而离子的转移过程需要转移蛋白参与,因而会有大量蛋白质分泌到胞外参与转移过程[40].其次是由于EPS中的蛋白质容易与金属离子通过静电作用而键合[41],降低细胞表面的负电性,促进污泥的絮凝作用,同时蛋白质中含有大量疏水基团,其含量的变化会改变微生物的表面性质,从而促进污泥的颗粒化进程.对比表 5各类颗粒污泥EPS组分及含量可发现,厌氧氨氧化颗粒污泥的EPS含量高于其他类型的颗粒污泥,说明厌氧氨氧化菌可分泌较多的EPS[13]. 表 5所列出的不同接种污泥形成的厌氧氨氧化颗粒污泥PN/PS差别较大,而同种接种污泥形成的厌氧氨氧化颗粒污泥PN/PS接近,因此推断接种污泥对厌氧氨氧化颗粒污泥PN/PS影响较大.
|
|
表 5 厌氧氨氧化颗粒污泥及其它种类颗粒污泥EPS组分 Table 5 EPS components in anammox granules and other different granules |
厌氧氨氧化颗粒污泥的多重荧光染色结果也证明了蛋白质是EPS的主要组分.如图 6(c)~6(e)所示,蛋白质的荧光强度和面积均大于多糖,说明蛋白质的含量高于多糖. β-D-呋喃葡萄糖和死细胞集中分布在颗粒污泥最外层,而活细胞、蛋白质、脂类、α-呋喃葡萄糖和α-甘露糖遍布整个颗粒污泥,并主要集中在外侧. McSwain等[11]认为非细胞类蛋白质负责维持颗粒污泥的稳定性,因此可以认为蛋白质和脂类构成了厌氧氨氧化颗粒污泥的骨架,厌氧氨氧化菌分布在蛋白质和脂类物质中间.多糖主要集中在颗粒污泥外层,可能是由于多糖包含大量的亲水性基团,如羧基,有利于厌氧氨氧化菌获得基质.其次亲水性的EPS可阻止气泡的相互黏附,从而避免颗粒污泥上浮现象[41].由图 6(c)所示β-多糖类似壳一样包裹在颗粒污泥最外层,是因为颗粒污泥外部不溶解性的β-多糖可增强颗粒污泥的稳定性[49]. Adav等[50]研究发现分布在颗粒外层的β-多糖对维持颗粒污泥稳定性的贡献大于内部的β-多糖. Tay等[51]发现,碳水化合物可促进细胞间的黏附,进而稳定微生物聚集体的结构.活细胞主要集中在外侧,颗粒污泥内部较少,这一结果与颗粒污泥的FISH检测结果一致,推测这可能与厌氧氨氧化颗粒污泥的形成过程有关.厌氧氨氧化菌在颗粒形成过程中以填充的形式进入颗粒内部,但外部基质充足内部基质较少,因此活细胞形成了这种独特的分布.死细胞集中在外侧可能是由于进水没有去除氧气,颗粒污泥外侧的厌氧氨氧化菌因接触氧气失活.

|
(a)明场;(b)c~f单个图像的叠加;(c)蓝色(Calcofluor white)β-D-呋喃葡萄糖;(d)青色(Con A)α-呋喃葡萄糖和α-甘露糖;(e)绿色(FITC)蛋白质;(f)黄色(尼罗红)脂类;(g)紫红色(SYTOX blue)死细胞;(h)红色(SYTO 63) 总细胞 图 6 厌氧氨氧化颗粒污泥的多重荧光染色(bar=40 μm) Fig. 6 Multiple fluorescent staining of ANAMMOXA granular sludge(bar=40 μm) |
采用ASBR反应器,接种硝化污泥快速启动了厌氧氨氧化反应.反应器稳定运行时,NH4+-N、NO2--N去除率均达到99%以上,TN去除率为89.87%±0.43%,总氮(TN)去除负荷达到1.7 kg·(m3·d)-1.本实验中厌氧氨氧化反应NH4+-N、NO2--N与NO3--N三者之间的比值为1:(1.32±0.08):(0.24±0.03).实验过程中发现,进出水pH差值、进水量(ΔpH×Q)与总氮负荷间存在线性相关性,出水NO3--N浓度同进水TN浓度间同样具有良好的线性相关性,因此在运行条件不变的前提下可通过出水pH和NO3--N浓度快速判断反应器运行情况. EPS含量的结果表明蛋白质为厌氧氨氧化颗粒污泥EPS的主要组分,蛋白质和多糖的含量分别为(59.61±5.64) mg·g-1、(12.21±2.04) mg·g-1,PN/PS为4.88±1.39.多重荧光染色的结果表明,β-D-呋喃葡萄糖和死细胞集中分布在颗粒污泥最外层;活细胞、蛋白质、脂类、α-呋喃葡萄糖和α-甘露糖遍布整个颗粒污泥,并主要集中在外侧.蛋白质和脂类构成了厌氧氨氧化颗粒污泥的骨架,厌氧氨氧化菌分布在蛋白质和脂类物质中间. β-D-呋喃葡萄糖包裹在厌氧氨氧化颗粒污泥最外层,对维持颗粒污泥结构稳定起重要作用.
| [1] | Bagchi S, Biswas R, Nandy T. Autotrophic ammonia removal processes:ecology to technology[J]. Critical Reviews in Environmental Science and Technology, 2012, 42(13): 1353–1418. DOI: 10.1080/10643389.2011.556885 |
| [2] | Kuenen J G. Anammox bacteria:from discovery to application[J]. Nature Reviews Microbiology, 2008, 6(4): 320–326. DOI: 10.1038/nrmicro1857 |
| [3] | Schmidt I, Sliekers O, Schmid M, et al. New concepts of microbial treatment processes for the nitrogen removal in wastewater[J]. FEMS Microbiology Reviews, 2003, 27(4): 481–492. DOI: 10.1016/S0168-6445(03)00039-1 |
| [4] | Terda A, Zhou S, Hosomi M. Presence and detection of anaerobic ammonium-oxidizing(anammox) bacteria and appraisal of anammox process for high-strength nitrogenous wastewater treatment:a review[J]. Clean Technologies and Environmental Policy, 2011, 13(6): 759–781. DOI: 10.1007/s10098-011-0355-3 |
| [5] | Henze M, van Loosdrecht M C M, Ekama G A, et al. Biological wastewater treatment:principles, modelling and design[M]. London: IWA Publishing, 2008: 139-154. |
| [6] | López H, Puig S, Ganigué R, et al. Start-up and enrichment of a granular anammox SBR to treat high nitrogen load wastewaters[J]. Journal of Chemical Technology and Biotechnology, 2008, 83(3): 233–241. DOI: 10.1002/(ISSN)1097-4660 |
| [7] | Chen T T, Zheng P, Shen L D. Growth and metabolism characteristics of anaerobic ammonium-oxidizing bacteria aggregates[J]. Applied Microbiology and Biotechnology, 2013, 97(12): 5575–5583. DOI: 10.1007/s00253-012-4346-z |
| [8] | Tang C J, Zheng P, Wang C H, et al. Performance of high-loaded ANAMMOX UASB reactors containing granular sludge[J]. Water Research, 2011, 45(1): 135–144. DOI: 10.1016/j.watres.2010.08.018 |
| [9] | Forster C F. Anaerobic upflow sludge blanket reactors:aspects of their microbiology and their chemistry[J]. Journal of Biotechnology, 1991, 17(3): 221–231. DOI: 10.1016/0168-1656(91)90013-L |
| [10] | Schmidt J E E, Ahring B K. Extracellular polymers in granular sludge from different upflow anaerobic sludge blanket(UASB) reactors[J]. Applied Microbiology and Biotechnology, 1994, 42(2-3): 457–462. |
| [11] | McSwain B S, Irvine R L, Hausner M, et al. Composition and distribution of extracellular polymeric substances in aerobic flocs and granular sludge[J]. Applied and Environmental Microbiology, 2005, 71(2): 1051–1057. DOI: 10.1128/AEM.71.2.1051-1057.2005 |
| [12] | Liu Y, Tay J H. The essential role of hydrodynamic shear force in the formation of biofilm and granular sludge[J]. Water Research, 2002, 36(7): 1653–1665. DOI: 10.1016/S0043-1354(01)00379-7 |
| [13] | Ni S Q, Fessehaie A, Lee P H, et al. Interaction of anammox bacteria and inactive methanogenic granules under high nitrogen selective pressure[J]. Bioresource Technology, 2010, 101(18): 6910–6915. DOI: 10.1016/j.biortech.2010.03.125 |
| [14] | 张丽丽, 陈效, 陈建孟, 等. 胞外多聚物在好氧颗粒污泥形成中的作用机制[J]. 环境科学, 2007, 28(4): 4795–4799. Zhang L L, Chen X, Chen J M, et al. Role mechanism of extracellular polymeric substances in the formation of aerobic granular sludge[J]. Environmental Science, 2007, 28(4): 4795–4799. |
| [15] | 王浩宇, 苏本生, 黄丹, 等. 好氧污泥颗粒化过程中Zeta电位与EPS的变化特性[J]. 环境科学, 2012, 33(5): 1614–1620. Wang H Y, Su B S, Huang D, et al. Profiles of Zeta potential and EPS in granulation process of aerobic sludge[J]. Environmental Science, 2012, 33(5): 1614–1620. |
| [16] | Li Y, Huang Z X, Ruan W Q, et al. ANAMMOX performance, granulation, and microbial response under COD disturbance[J]. Journal of Chemical Technology and Biotechnology, 2015, 90(1): 139–148. DOI: 10.1002/jctb.2015.90.issue-1 |
| [17] | 宋成康, 王亚宜, 韩海成, 等. 温度降低对厌氧氨氧化脱氮效能及污泥胞外聚合物的影响[J]. 中国环境科学, 2016, 36(7): 2006–2013. Song C K, Wang Y Y, Han H C, et al. Effect of decreasing temperature on the performance and extracellular polymer substance of anaerobic ammonia oxidation sludge[J]. China Environmental Science, 2016, 36(7): 2006–2013. |
| [18] | van de Graaf A A, de Bruijn P, Robertson L A, et al. Autotrophic growth of anaerobic ammonium-oxidizing micro-organisms in a fluidized bed reactor[J]. Microbiology, 1996, 142(8): 2187–2196. DOI: 10.1099/13500872-142-8-2187 |
| [19] | Amann R I, Binder B J, Olson R J, et al. Combination of 16S rRNA-targeted oligonucleotide probes with flow cytometry for analyzing mixed microbial populations[J]. Applied and Environmental Microbiology, 1990, 56(6): 1919–1925. |
| [20] | Daims H, Brühl A, Amann R, et al. The domain-specific probe EUB338 is insufficient for the detection of all bacteria:development and evaluation of a more comprehensive probe set[J]. Systematic and Applied Microbiology, 1999, 22(3): 434–444. DOI: 10.1016/S0723-2020(99)80053-8 |
| [21] | Schmid M, Walsh K, Webb R, et al. Candidatus "Scalindua brodae", sp. nov., Candidatus "Scalindua wagneri", sp. nov., two new species of anaerobic ammonium oxidizing bacteria[J]. Systematic and Applied Microbiology, 2003, 26(4): 529–538. DOI: 10.1078/072320203770865837 |
| [22] | Frølund B, Palmgren R, Keiding K, et al. Extraction of extracellular polymers from activated sludge using a cation exchange resin[J]. Water Research, 1996, 30(8): 1749–1758. DOI: 10.1016/0043-1354(95)00323-1 |
| [23] | DuBois M, Gilles K A, Hamilton J K, et al. Colorimetric method for determination of sugars and related substances[J]. Analytical Chemistry, 1956, 28(3): 350–356. DOI: 10.1021/ac60111a017 |
| [24] | Chen M Y, Lee D J, Tay J H, et al. Staining of extracellular polymeric substances and cells in bioaggregates[J]. Applied Microbiology and Biotechnology, 2007, 75(2): 467–474. DOI: 10.1007/s00253-006-0816-5 |
| [25] | 路青, 华兆哲, 李秀芬, 等. MBR中厌氧氨氧化运行特性及微生态结构[J]. 食品与生物技术学报, 2010, 29(4): 581–588. Lu Q, Hua Z Z, Li X F, et al. Operation characterictics and microbial community distribution of anammox in a membrane bioreactor[J]. Journal of Food Science and Biotechnology, 2010, 29(4): 581–588. |
| [26] | Tang C J, Zheng P, Mahmood Q, et al. Start-up and inhibition analysis of the anammox process seeded with anaerobic granular sludge[J]. Journal of Industrial Microbiology & Biotechnology, 2009, 36(8): 1093–1100. |
| [27] | Wang T, Zhang H M, Gao D W, et al. Enrichment of anammox bacteria in seed sludges from different wastewater treating processes and start-up of anammox process[J]. Desalination, 2011, 271(1-3): 193–198. DOI: 10.1016/j.desal.2010.12.034 |
| [28] | van der Star W R L, Abma W R, Blommers D, et al. Startup of reactors for anoxic ammonium oxidation:experiences from the first full-scale anammox reactor in rotterdam[J]. Water Research, 2007, 41(18): 4149–4163. DOI: 10.1016/j.watres.2007.03.044 |
| [29] | Zhang L, Yang J C, Ma Y G, et al. Treatment capability of an up-flow anammox column reactor using polyethylene sponge strips as biomass carrier[J]. Journal of Bioscience and Bioengineering, 2010, 110(1): 72–78. DOI: 10.1016/j.jbiosc.2009.12.005 |
| [30] | Liu S T, Yang F L, Gong Z, et al. Application of anaerobic ammonium-oxidizing consortium to achieve completely autotrophic ammonium and sulfate removal[J]. Bioresource Technology, 2008, 99(15): 6817–6825. DOI: 10.1016/j.biortech.2008.01.054 |
| [31] | Li H S, Zhou Q S, Ma W H, et al. Fast start-up of ANAMMOX reactor:operational strategy and some characteristics as indicators of reactor performance[J]. Desalination, 2012, 286: 436–441. DOI: 10.1016/j.desal.2011.11.038 |
| [32] | Strous M, Heijnen J J, Kuenen J G, et al. The sequencing batch reactor as a powerful tool for the study of slowly growing anaerobic ammonium-oxidizing microorganisms[J]. Applied Microbiology and Biotechnology, 1998, 50(5): 589–596. DOI: 10.1007/s002530051340 |
| [33] | 冯平, 周少奇. 常温下厌氧氨氧化生物膜反应器的启动研究[J]. 环境科学与技术, 2010, 33(6): 19–22. Feng P, Zhou S Q. Experimental study on start-up of UASB-anammox biofilm reactor at ordinary temperature[J]. Environmental Science & Technology, 2010, 33(6): 19–22. |
| [34] | 张海芹, 王翻翻, 李月寒, 等. 不同接种污泥ABR厌氧氨氧化的启动特征[J]. 环境科学, 2015, 36(6): 2216–2221. Zhang H Q, Wang F F, Li Y H, et al. Start-up performance of ANAMMOX enrichment with different inoculated sludge in anaerobic baffled reactor[J]. Environmental Science, 2015, 36(6): 2216–2221. |
| [35] | Ahn Y H, Hwang I S, Min K S. ANAMMOX and partial denitritation in anaerobic nitrogen removal from piggery waste[J]. Water Science & Technology, 2004, 49(5-6): 145–153. |
| [36] | Ahn Y H. Sustainable nitrogen elimination biotechnologies:a review[J]. Process Biochemistry, 2006, 41(8): 1709–1721. DOI: 10.1016/j.procbio.2006.03.033 |
| [37] | Araujo J C, Campos A C, Correa M M, et al. Anammox bacteria enrichment and characterization from municipal activated sludge[J]. Water Science & Technology, 2011, 64(7): 1428–1434. |
| [38] | 赵弋戈, 郑平. 厌氧氨氧化体的组成、结构与功能[J]. 微生物学报, 2016, 56(1): 8–18. Zhao Y G, Zheng P. Composition, structure and function of anammoxosome-a review[J]. Acta Microbiologica Sinica, 2016, 56(1): 8–18. |
| [39] | 杨敏, 胡学伟, 宁平, 等. 废水生物处理中胞外聚合物(EPS)的研究进展[J]. 工业水处理, 2011, 31(7): 7–12. Yang M, Hu X W, Ning P, et al. Research progress in extracellular polymeric substances applied to biological wastewater treatment[J]. Industrial Water Treatment, 2011, 31(7): 7–12. DOI: 10.11894/1005-829x.2011.31(7).7 |
| [40] | 王然登, 程战利, 彭永臻, 等. 强化生物除磷系统中胞外聚合物的特性[J]. 中国环境科学, 2014, 34(11): 2838–2843. Wang R D, Cheng Z L, Peng Y Z, et al. Characteristics of EPS taken from an enhanced biological phosphorus removal system[J]. China Environmental Science, 2014, 34(11): 2838–2843. |
| [41] | Zhu L, Lv M L, Dai X, et al. Role and significance of extracellular polymeric substances on the property of aerobic granule[J]. Bioresource Technology, 2012, 107: 46–54. DOI: 10.1016/j.biortech.2011.12.008 |
| [42] | Ni S Q, Sun N, Yang H L, et al. Distribution of extracellular polymeric substances in anammox granules and their important roles during anammox granulation[J]. Biochemical Engineering Journal, 2015, 101: 126–133. DOI: 10.1016/j.bej.2015.05.014 |
| [43] | Ni B J, Hu B L, Fang F, et al. Microbial and physicochemical characteristics of compact anaerobic ammonium-oxidizing granules in an upflow anaerobic sludge blanket reactor[J]. Applied and Environment Microbiology, 2010, 76(8): 2652–2656. DOI: 10.1128/AEM.02271-09 |
| [44] | Li Z L, Xu X C, Shao B, et al. Anammox granules formation and performance in a submerged anaerobic membrane bioreactor[J]. Chemical Engineering Journal, 2014, 254: 9–16. DOI: 10.1016/j.cej.2014.04.068 |
| [45] | Jemaat Z, Suárez-Ojeda M E, Pérez J, et al. Partial nitritation and o-cresol removal with aerobic granular biomass in a continuous airlift reactor[J]. Water Research, 2014, 48: 354–362. DOI: 10.1016/j.watres.2013.09.048 |
| [46] | Wu J, Zhou H M, Li H Z, et al. Impacts of hydrodynamic shear force on nucleation of flocculent sludge in anaerobic reactor[J]. Water Research, 2009, 43(12): 3029–3036. DOI: 10.1016/j.watres.2009.04.026 |
| [47] | Martínez F, Lema J, Méndez R, et al. Role of exopolymeric protein on the settleability of nitrifying sludges[J]. Bioresource Technology, 2004, 94(1): 43–48. DOI: 10.1016/j.biortech.2003.11.010 |
| [48] | Bhatti Z I, Sumida K, Rouse J D, et al. Characterization of denitrifying granular sludge treating soft groundwater in an upflow sludge-blanket reactor[J]. Journal of Bioscience and Bioengineering, 2001, 91(4): 373–377. DOI: 10.1016/S1389-1723(01)80154-7 |
| [49] | Wang Z W, Liu Y, Tay J H. Distribution of EPS and cell surface hydrophobicity in aerobic granules[J]. Applied Microbiology and Biotechnology, 2005, 69(4): 469–473. DOI: 10.1007/s00253-005-1991-5 |
| [50] | Adav S S, Lee D J, Tay J H. Extracellular polymeric substances and structural stability of aerobic granule[J]. Water Research, 2008, 42(6-7): 1644–1650. DOI: 10.1016/j.watres.2007.10.013 |
| [51] | Tay J H, Liu Q S, Liu Y. The role of cellular polysaccharides in the formation and stability of aerobic granules[J]. Letters in Applied Microbiology, 2001, 33(3): 222–226. DOI: 10.1046/j.1472-765x.2001.00986.x |