 2017, Vol. 38
2017, Vol. 38



2. 哈尔滨工业大学城市水资源与水环境国家重点实验室, 哈尔滨 150090;
3. 江苏高校水处理技术与材料协同创新中心, 苏州 215009;
4. 宁波市城市排水有限公司, 宁波 315000;
5. 大化集团有限责任公司, 大连 116031
2. State Key Laboratory of Urban Water Resource and Environment, Harbin Institute of Technology, Harbin 150090, China;
3. Jiangsu College of Water Treatment Technology and Material Collaborative Innovation Center, Suzhou 215009, China;
4. Ningbo Municipal Drainage Co. Ltd., Ningbo 315000, China;
5. Dahua Group Co., Ltd., Dalian 116031, China
目前污水处理厂运行中存在出水总氮难以达标的现实问题, 且其去除受低温, 有机负荷和工业废水影响较大, 是污水处理厂稳定运行所面对的巨大挑战.对此, 污水处理厂一般采用投加碳源和悬浮填料的方式以增强总氮去除效果.在外加碳源充足的条件下, 将悬浮填料从投加到挂好缺氧生物膜至少需要11周时间[1], 周期相对较长, 而且污水处理厂缺氧单元需要投加的填料数目相当巨大, 无法满足短期内污水处理厂总氮迅速达标排放的要求.因此, 如果预先培养出大量缺氧生物膜填料, 当需要填料时只要将其取出, 经过短时间的活性恢复即可投入生产, 可以有效解决总氮无法稳定达标排放的现实问题, 同时可以提高污水处理厂生产效率.
投加悬浮填料到污水处理厂操作单元过程中可能遇到各种不同环境条件, 如南方城市降水较多, 排水体制采取合流制方式, 排水管网渗漏[2], 均会对城镇污水产生稀释作用, 导致进水有机负荷明显降低; 上游工业废水水质波动较大, 进水存在冲击负荷; 冬季温度较低导致微生物代谢活性下降等, 这些条件都会对填料的活性恢复效果产生影响.因此, 研究悬浮填料活性恢复过程在低负荷, 冲击负荷及低温条件的适用性, 对悬浮填料储存与活性恢复技术在实际工程中能否开展广泛应用具有重要的理论和现实意义.
环境微生物群落是由大量的具有不同遗传性状的微生物组成.了解群落组成成分的变化有助于从宏观水平上认识群落的结构变化, 从而帮助解析生理生化特性产生变化的原因.随着分子生物学技术在污水生物处理领域的广泛应用, 人们对环境微生物有了更多的认识.王昊昱等[3]利用454高通量测序法对厌氧消化反应器污泥中的细菌及古细菌进行了群落结构分析. Martín等[4]对Ca. Accumulibacter进行全基因组测序, 检测到了亚硝氮还原至氮气的基因.分子生物学技术已成为研究环境样品微生物群落结构的重要手段.
目前国内外的相关研究主要集中于填料材料开发[5, 6]、污染物去除特性[7, 8]及强化硝化反硝化[9~11]等, 而对于缺氧生物膜填料的活性恢复及其适用性的研究相对较少.因此, 本文将已完成缺氧挂膜的悬浮填料作为研究对象, 以正常负荷及常温下运行为对照, 考察不同恢复环境(低负荷、冲击负荷、低温)条件下缺氧生物膜填料在活性恢复过程中物理特性、污染物去除过程及反硝化性能, 解析悬浮填料反硝化活性恢复过程中微生物菌群结构, 以期为缺氧悬浮填料技术在实际工程中的应用提供理论依据和技术支持.
1 材料与方法 1.1 实验装置反应器由4个体积为5 L的有机玻璃容器组成, 顶部设有搅拌装置, 保证反应器内悬浮填料充分流化.每个反应器分别填充等量悬浮填料, 填充率为30%(体积分数), 实验装置如图 1所示.

|
图 1 实验装置示意 Fig. 1 Schematic diagram of experimental set-up |
取缺氧挂膜悬浮填料若干, 均分后分别置于4个反硝化反应器中, 将反应器分别按如下情况编号并设置实验条件. R1:对照组、R2:低负荷组、R3:冲击负荷组、R4:低温组, 具体进水水质及温度条件如表 1所示.实验均采用人工配水间歇进出水的方式, 水力停留时间为8 h.
|
|
表 1 人工配水水质 Table 1 Components and concentrations of synthetic wastewater |
1.3 分析指标与方法
(1) 水质分析实验过程中各项水质指标采用文献[12]的方法测定.
(2) 附着生物量测定实验用SPR-1型悬浮填料的材质为聚乙烯, 温度超过130℃时易变形分解, 难以用常规灼烧法测其生物量.故本实验结合Douterelo等[13]的测量方法并加以改进, 采用低温烘干的方式进行填料有效生物量的测定.具体步骤如下.
附着总生物膜量(TS, 单位:以TS/载体计, mg·g-1)测定:TS=(m1-m2)/取出填料总重量.从反应器中取出3~5个填料在70~80℃条件下烘干至恒重, 称重记m1; 用移液管刷将填料表面生物膜刷至烧杯中, 并用1 mg·L-1 NaOH浸泡填料并用电磁炉煮沸并搅拌60 min[11], 取出超声10 min, 用蒸馏水冲洗后烘干至恒重, 称重记为m2.
附着生物量(VS, 单位:以VS/载体计, mg·g-1)测定:
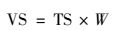
|
式中, W:将填料表面刷下的生物膜置于坩埚内, 于105℃烘箱内烘干称重m3, 再于500℃马弗炉内灼烧后称重m4及干锅重量m0, 则W=(m4-m0)/(m3-m0).
(3) 生物膜EPS提取与测量EPS是微生物代谢产生的位于细胞表面的高分子聚合物质, 本实验采用甲酰胺-NaOH的方法提取生物膜的EPS[14], 通过测试生物膜EPS中蛋白质和多糖的含量来反映生物膜EPS含量, 样品采用蒽酮比色法测定多糖类物质的含量[15], 考马斯亮蓝法测定蛋白质类物质的含量[16].
(4) 填料微生物菌群分子生物学分析取若干填料样品, 用超纯水清洗后将生物膜刷于灭菌的Eppendorf离心管中, 加入2 mL 1×PBS混匀, 在5 000 r·min-1条件下离心5 min, 弃去上清液, 如此清洗生物膜样品2~3次.采用ABigen土壤DNA提取试剂盒(柱形)提取填料微生物总宏基因组, 然后用1%琼脂糖凝胶电泳(电压120 V, 时间15 min)来检测所提取DNA的浓度和纯度, 最后置于-20℃下保存备用. PCR扩增及HiSeq建库委托上海晶能生物技术有限公司进行, 测序平台为Illumina HiSeq, 采用Trimmomatic软件对序列进行去杂, 采用Qiime软件进行群落丰度计算.
2 结果与讨论 2.1 附着生物量变化如图 2所示, 4组反应器的悬浮填料在活性恢复初期附着生物量均呈下降趋势, 原因是填料在储存过程中缺乏营养物质, 部分生物膜失活脱落.随后R1~R4生物量均有所恢复, 最大值分别为93.79、53.39、86.14和77.82 mg·g-1.其中前5 d内R2和R4生物量下降更多, 表明低温及低负荷条件对微生物活性恢复初期影响更大.但随着恢复时间的增加, R4生物量快速升高, 说明填料附着微生物能够进行自我调节, 提高自身对低温的适应性.这种自我调节可能是通过改变自身代谢途径, 建立一种新的低温代谢和种群结构来实现的[18].而R2生物量增加缓慢, 至恢复周期结束其生物量仅比恢复过程启动时提高了16.69%, 表明微生物生长代谢受进水有机负荷限制, 生物膜增殖速率缓慢, 在长期饥饿条件下进行内源呼吸作用不利于其生物活性的恢复.因此在低负荷条件下, 缺氧挂膜填料不适用于短期降低出水总氮的情况.

|
图 2 不同反应器填料附着生物量变化 Fig. 2 Biomass variations of suspended carriers of R1-R4 |
不同环境下微生物分泌的胞外聚合物化学组成不同, 通过影响细胞间的黏附过程和生物膜内部结构对悬浮填料活性恢复起到重要作用[19].由图 3可以看出, 在恢复过程前5 d 4组反应器填料生物膜溶解性EPS中的蛋白质和多糖含量均呈明显下降趋势.这是由于储存过程中营养物质和溶解氧缺乏, 填料生物膜外层微生物处于饥饿条件下, 会利用胞外聚合物以保持其生存能力和微生物活性[20].反应器开始运行后, 外层微生物活性迅速恢复, 为了维持生物膜内外微生物平衡, 生物膜内部溶解性EPS被转化用来维持生物膜完整的结构.生物膜溶解性EPS中的蛋白质和多糖含量与图 2中R1~R4填料附着生物量前5 d变化趋势一致, 与McSwain等[21]所得的附着生物膜量与EPS(即蛋白质和多糖)存在密切联系的研究结论相符. R1中填料微生物对多糖的合成速率高于蛋白质, 随着恢复时间的增加, 生物膜性质、多糖和蛋白质类物质浓度均趋于稳定.有研究表明[22, 23], 生物膜EPS中的蛋白质能够强化相邻细胞间相互交联作用, 较高的PN浓度有利于维持生物膜内部结构和生物量, 而较高的PS浓度能够有效提高细胞的黏附作用, 维持生物膜外层结构的稳定.本阶段采用的悬浮填料在储存之前已经完成缺氧挂膜, 其内部生物膜结构比较完整, 而细胞间黏附性在储存过程中减弱, 细胞膜外层结构遭到一定程度的破坏, 因此在活性恢复过程中起关键作用的是EPS中多糖的增加.

|
图 3 生物膜EPS中蛋白质和多糖含量的变化 Fig. 3 Changes in protein(PN) and polysaccharides(PS) contents of biofilm EPS during reactivation |
R3除了在受到负荷冲击时多糖含量有所波动之外, 蛋白质含量及正常负荷状态下的多糖含量变化趋势与R1基本相同, 这是因为在有机负荷较高时, 微生物不能充分利用过量的碳源, 而将其转变为多糖类物质储存在胞外[24], 使EPS中多糖含量升高.而R2中多糖和蛋白质含量均比R1高, 其原因是在高负荷条件能够满足微生物代谢所需的营养物质, 因此微生物分泌的EPS含量相对稳定, 而低负荷意味着进水中营养物质较少, 微生物有较慢的生长速率和较高的内源代谢水平, 部分细胞发生自溶, 使EPS含量增加[25]. R4的多糖含量增加比蛋白质更为明显, 这是由于温度影响细胞的生理活动, 随着温度的降低, 酶促反应速度会降低[26], 细胞分泌蛋白质的过程受到一定抑制.
2.3 污染物去除特性 2.3.1 NO3--N的还原由图 4可知, 随着恢复时间的增加, 各反应器中填料对NO3--N的还原率均有所上升, 其中R1在3 d时达到90%以上, R4则在5 d时恢复到同等水平, 并在之后的时间里分别保持平均95.89%和95.18%的稳定NO3--N还原能力, 这与2.1节中R4在5 d后附着生物量快速升高相吻合. R3在3 d时NO3--N还原率达到95.58%后, 无论有机负荷如何变化, 出水中硝酸盐始终保持很低的浓度, NO3--N还原率维持在90%以上, 这表明较高的碳源浓度对硝酸盐的还原不会产生较大影响, 冲击负荷不会干扰悬浮填料微生物硝酸盐还原能力的恢复.相比之下, R2的NO3--N还原率在3 d时仅为56.4%, 在悬浮填料恢复过程中, 其NO3--N还原率最高仅为74.36%, 在25 d的恢复过程中反硝化一直不完全, 与周娜[18]的研究结果较为一致.主要原因在于微生物在低负荷下长时间处于内源代谢期, 碳源首先部分贮存在体内以保持菌体生长, 导致用于反硝化过程的碳源浓度进一步降低[27].但与活性污泥系统在低负荷条件下59.4%的NO3--N平均还原率[28]相比, 悬浮填料系统(NO3--N平均还原率为67.8%)更适用于低负荷条件.

|
图 4 各反应器对NO3--N的还原率 Fig. 4 NO3--N reductive efficiencies of R1-R4 during the reactivation |
亚硝酸盐氮(NO2--N)是反硝化过程的中间产物, 其积累会对反硝化菌产生毒害作用, 抑制反硝化过程, 进而对污水处理厂生物处理工艺的正常运行造成破坏.此外, 在常规脱氮工艺中, 积累有大量NO2--N的废水离开缺氧池后会在之后的好氧池中重新被氧化为NO3--N, 使得脱氮时需氧量增大, 不但降低处理效果, 而且浪费了能源[29].
4组反应器在活性恢复过程的NO2--N质量浓度变化如图 5所示, R1和R3仅在前2 d出现了NO2--N积累现象, 之后出水NO2--N质量浓度均低于0.03 mg·L-1. R2前2 d出水NO2--N较低, 之后出现了严重的NO2--N积累, 到12 d之后出水中才没有NO2--N检出. R2中前2 d和12 d之后NO2--N质量浓度均较低但原因不同:在亚硝酸盐氮积累阶段, 同时存在亚硝酸盐氮的增加(硝酸盐氮还原产生)和减少(亚硝酸盐氮还原为氮气), 可认为亚硝酸盐氮积累是反硝化过程中两步反应速率差造成的[30], 前2 d内R2中硝酸盐还原菌和亚硝酸盐还原菌活性均未恢复, 结合图 4中前2 d内NO3--N还原率较低, NO3--N还原产生的NO2--N本就较少, 因此出水NO2--N质量浓度低; 而12 d后随着硝酸盐还原菌和亚硝酸盐还原菌活性缓慢恢复, NO3--N还原产生的NO2--N均可以被进一步还原, 因此没有出现NO2--N积累现象. R2恢复前期NO2--N积累严重是由于在低负荷条件下碳源缺乏所致, 与硝酸盐还原菌相比, 亚硝酸盐还原菌在争夺电子受体时处于劣势[31].而R4恢复前期同样出现了NO2--N严重积累, 这是由于亚硝酸盐还原菌比硝酸盐还原菌对温度更为敏感, 受到低温抑制作用更大.虽原因不同, 但R2和R4两个反应器的亚硝酸盐还原菌活性恢复都受到了影响, 导致硝酸盐还原为亚硝酸盐的速率大于亚硝酸盐还原为氮气的速率, 表现出亚硝酸盐的积累.经过时间不等的活性恢复过程后, 4个反应器的缺氧挂膜悬浮填料生物膜均已具有良好而稳定的降解亚硝酸盐的能力.

|
图 5 各反应器NO2--N质量浓度的变化 Fig. 5 Changes of NO2--N mass concentration of R1-R4 during the reactivation |
由图 6可以看出, 4个反应器的反硝化效能均随恢复时间的增加有所恢复, 其中R1和R3的恢复效果较明显, 运行到5 d时反硝化速率(以NO3--N/VSS计)均达到为1.5 mg·(g·h)-1左右, R4达到相同水平需要10 d. R1、R3和R4反硝化速率均在15 d后逐渐恢复稳定, 数值为5.4 mg·(g·h)-1左右, 且均在160 min左右将NO3--N质量浓度降至1 mg·L-1以下.而R2在25 d的活性恢复过程中均不能在300 min内实现完全反硝化, NO3--N还原率仅由46.68%提高到64.71%.这是由于在低负荷状态下, 反硝化异养菌将分别利用易降解有机物和内碳源, 导致其反硝化能力大幅下降[32].

|
图 6 不同反应器硝酸盐还原特性 Fig. 6 Characteristics of nitrate reduction of R1-R4 |
悬浮填料的脱氮过程包括硝酸盐氮还原为亚硝酸盐氮和亚硝酸盐氮还原为氮气两个步骤, 因此评价其脱氮能力应当同时考虑硝酸盐的还原和亚硝酸盐的积累两个效果.如图 7所示, R1及R3运行到10 d时即可实现300 min内NO2--N的全部还原, 并且这一时间在15 d后减少为180 min, R4达到相同亚硝酸盐零积累能力分别需15 d和20 d.而R2运行到20 d时, 300 min内依然有0.053 mg·L-1的NO2--N积累, 25 d才可以用250 min的时间完成亚硝态氮零积累.以上结果表明, 缺氧悬浮填料的活性恢复对冲击负荷条件的适用性较好, 低温条件次之, 对低负荷环境适用性较差.因此当应用缺氧悬浮填料的污水处理厂存在处理水质负荷较低的问题时, 需要考虑通过额外投加碳源的方式提高脱氮能力.

|
图 7 不同反应器亚硝酸盐积累特性 Fig. 7 Characteristics of nitrite accumulation in R1-R4 |
本实验采用Illumina HiSeq 3000测序平台对各反应器内不同恢复时期的悬浮填料缺氧生物膜进行16S rRNA高通量测序, 以表征恢复过程中的生物膜微生物群落结构变化.根据测序报告中丰富度稀疏分析图的曲线趋于平缓可得出, 本实验测序数据量合理, 更多的数据只会产生少量新的操作分类单元, 即所获实验测序数据准确可靠.
采用Chao1指数来分析不同运行阶段的缺氧生物膜微生物多样性的物种丰富度, 用香农指数(Shannon)来分析其均匀度与丰富度综合特征(图 8).运行第15 d时, R2反应器微生物Shannon指数和Chao1指数分别为3.67和385, 均高于其他反应器.可知低负荷下的缺氧挂膜悬浮填料的生物膜中物种较丰富, 但由于其与氮类污染物还原有关的菌种含量较低, 此时R2反应器的脱氮效果不理想.当运行到25 d时, 各组反应器的Shannon指数和Chao1指数基本相同, 表明不同恢复环境对缺氧生物膜恢复完成时的生物多样性水平影响较小.

|
图 8 种群生物多样性指标变化 Fig. 8 Changes of community biodiversity index |
由图 9可知, 不同运行时期缺氧填料生物膜中, Proteobacteria菌门和Bacteroidetes菌门是最占优势的两个细菌菌门.生物膜中Proteobacteria菌门的相对丰度随着培养时间的延长除在R1中略微下降外, 在R2、R3和R4反应器中均逐渐升高, 分别由15 d时的72.23%、78.66%和76.35%上升至25 d时的83.17%、84.30%和80.46%;Bacteroidetes菌门除在R1中稍有上升外, 在R2、R3和R4反应器中均呈相对下降的趋势, 分别由15 d时的13.46%、19.56%和32.57%下降至25 d时的7.63%、17.64%和13.23%.对测序结果进一步分析可知, Proteobacteria菌门中主要组分为β-Proteobacteria, 研究表明, β-Proteobacteria广泛存在于污水处理过程中, 是降解氮类污染物的重要菌群, 其中有大部分菌属能够在厌/缺氧条件下降解有机物污染物进行生长和繁殖[33]. R2的生物膜在第15 d时Proteobacteria菌相对丰度低于其他反应器, 表明短期内低负荷下填料脱氮效果恢复不佳, 不能作为污水处理厂低负荷条件下总氮快速达标的备选方案.

|
图 9 填料生物膜细菌菌门分布 Fig. 9 Distributions of bacterial strains at level of phylum |
进一步将微生物菌群以属划分, 结果如图 10所示.在8个样品中相对丰度最高的细菌菌属有:Dechloromonas、Simplicispira、Pseudomonas、Flavobacterium、Zoogloea和Ferribacterium, 其相对丰度之和分别占8个样品总菌属的83.16%、71.83%、49.02%、72.66%、46.36%、58.99%、60.22%和72.53%.随着恢复时间的增加, 填料生物膜中的细菌种类均出现了较大变化.对照组R1反应器中生物膜微生物菌属中所占比例最大的为Dechloromonas菌属, 占总菌属的40%以上, R2中Dechloromonas菌属相对丰度仅为10%左右. R3中最优势的菌种除Dechloromonas菌属外还有Pseudomonas菌属, 且这两种菌属相对丰度之和由15 d时的33.38%增加到25 d时的46.92%.有研究表明, Dechloromonas菌属是一类典型的具有反硝化功能的菌属[34], 其中典型的菌种如D. agitate和D. denitrificans都具有一定的硝酸盐降解能力[35], 而Pseudomonas菌属具有较高的脱氮除磷特性[36].这与R1和R3反应器脱氮效果较好的现象符合.与对照组R1进行比较, R4中Dechloromonas菌属的相对丰度由15 d时的1.09%增加到25 d时的35.47%, Pseudomonas菌属则由41.91%下降到12.84%, 而R4反应器依然能够较快且较好地恢复脱氮效果, 因此反硝化功能菌属Dechloromonas相对丰度的增加可能是引起填料生物膜反硝化脱氮能力提高的重要原因.另外, R4反应器Flavobacterium菌属的相对丰度在15 d和25 d时分别为14.30%和9.48%, 均高于同时期的R1~R3反应器.有研究表明, Flavobacterium菌属能够还原硝酸盐且适合在低温下生长[37], 因此Flavobacterium菌属的存在可能在一定程度上促进了低温条件下填料生物膜反硝化脱氮能力的恢复.

|
图 10 填料生物膜细菌菌属分布 Fig. 10 Distributions of bacterial strains at level of genus |
(1) 冲击负荷对附着生物量恢复影响不大, 低温及低负荷条件对微生物活性恢复初期影响较大.但随着恢复时间的增加, 填料附着微生物能够进行自我调节, 提高自身对低温的适应性.低负荷下生物膜增殖速率缓慢, 不利于其生物活性的恢复.
(2) 在填料活性恢复过程中起关键作用的是EPS中多糖的增加.冲击负荷和低温条件下多糖含量增加比蛋白质更为明显.而低负荷下由于部分细胞自溶, 导致多糖和蛋白质含量比另外两者要多.
(3) 挂膜填料脱氮效能恢复在一定的负荷冲击范围内不会受其影响, 低温条件对填料脱氮能力恢复的影响较小, 低负荷下脱氮能力恢复有限.因此缺氧挂膜悬浮填料的活性恢复对冲击负荷条件的适用性较好, 低温条件次之, 对低负荷环境适用性较差.
(4) 缺氧悬浮填料不能作为污水处理厂低负荷下总氮快速达标的备选方案.反硝化功能菌属Dechloromonas相对丰度的增加可能是引起填料生物膜反硝化脱氮能力提高的重要原因. Flavobacterium菌属的存在可能促进了低温下填料生物膜反硝化脱氮能力的恢复.不同恢复环境对缺氧生物膜恢复完成时的生物多样性影响较小.
| [1] | 任彦强, 赵雪莲, 李宗慧, 等. MBBR悬浮填料低温处理生活污水对比实验研究[J]. 河北工业科技, 2014, 31(3): 215–219. Ren Y Q, Zhao X L, Li Z H, et al. Comparative research on MBBR suspended carriers for the municipal sewage treatment at low temperature[J]. Hebei Journal of Industrial Science and Technology, 2014, 31(3): 215–219. |
| [2] | 温荣平, 卢东昱, 郭伟, 等. 低负荷城市污水厂的运行问题与对策[J]. 中国给水排水, 2010, 26(18): 144–146. Wen R P, Lu D Y, Guo W, et al. Operation problems and countermeasures of municipal wastewater treatment plant at low organic loading[J]. China Water & Wastewater, 2010, 26(18): 144–146. |
| [3] | 王昊昱, 陶彧, 任南琪. 底物类型对产甲烷效能及微生物群落结构的影响[J]. 哈尔滨工业大学学报, 2016, 48(2): 9–14. Wang H Y, Tao Y, Ren N Q. Impact of organic matter type on the efficiency and microbial community structure of an anaerobic digestion process[J]. Journal of Harbin Institute of Technology, 2016, 48(2): 9–14. DOI: 10.11918/j.issn.0367-6234.2016.02.002 |
| [4] | Martín H G, Ivanova N, Kunin V, et al. Metagenomic analysis of two enhanced biological phosphorus removal (EBPR) sludge communities[J]. Nature Biotechnology, 2006, 24(10): 1263–1269. DOI: 10.1038/nbt1247 |
| [5] | 海景, 黄尚东, 张凡, 等. 改性聚丙烯生物填料的制备与应用[J]. 化工环保, 2006, 26(4): 333–336. Hai J, Huang S D, Zhang F, et al. Preparation of modified polypropylene biofilm carrier and its application[J]. Environmental Protection of Chemical Industry, 2006, 26(4): 333–336. |
| [6] | Ferrai M, Guglielmi G, Andreottola G. Modelling respirometric tests for the assessment of kinetic and stoichiometric parameters on MBBR biofilm for municipal wastewater treatment[J]. Environmental Modelling & Software, 2010, 25(5): 626–632. |
| [7] | 荆洁颖. 移动床生物膜反应器对焦化废水脱碳除氮研究[D]. 太原: 太原理工大学, 2009. |
| [8] | Majone M, Aulenta F, Dionisi D, et al. High-rate anaerobic treatment of Fischer-Tropsch wastewater in a packed-bed biofilm reactor[J]. Water Research, 2010, 44(9): 2745–2752. DOI: 10.1016/j.watres.2010.02.008 |
| [9] | 郑敏, 杨波, 汪诚文, 等. 中试MBBR装置强化氨氮去除速率的影响条件研究[J]. 中国环境科学, 2012, 32(10): 1778–1783. Zheng M, Yang B, Wang C W, et al. Enhanced ammonia removal rate in a pilot-scale MBBR[J]. China Environmental Science, 2012, 32(10): 1778–1783. DOI: 10.3969/j.issn.1000-6923.2012.10.008 |
| [10] | 郑兰香, 鞠兴华. 温度和C/N比对生物膜反硝化速率的影响[J]. 西南给排水, 2005, 27(2): 31–33. Zheng L X, Ju X H. The effect of temperature and C/N radio on biofilm denitirification rate[J]. Southwest Water and Wastewater, 2005, 27(2): 31–33. |
| [11] | Mases M, Dimitrova I, Nyberg U, et al. Experiences from MBBR post-denitrification process in long-term operation at two WWTPs[J]. Proceedings of the Water Environment Federation, 2010, 2010(7): 458–471. DOI: 10.2175/193864710798208791 |
| [12] | 国家环境保护总局. 水和废水监测分析方法[M]. (第四版). 北京: 中国环境科学出版社, 2002: 210-276. |
| [13] | Douterelo I, Boxall J B, Deines P, et al. Methodological approaches for studying the microbial ecology of drinking water distribution systems[J]. Water Research, 2014, 65: 134–156. DOI: 10.1016/j.watres.2014.07.008 |
| [14] | Liang Z W, Li W H, Yang S Y, et al. Extraction and structural characteristics of extracellular polymeric substances (EPS), pellets in autotrophic nitrifying biofilm and activated sludge[J]. Chemosphere, 2010, 81(5): 626–632. DOI: 10.1016/j.chemosphere.2010.03.043 |
| [15] | Adav S S, Lee D J. Extraction of extracellular polymeric substances from aerobic granule with compact interior structure[J]. Journal of Hazardous Materials, 2008, 154(1-3): 1120–1126. DOI: 10.1016/j.jhazmat.2007.11.058 |
| [16] | 刘美, 王湛. 胞外聚合物对膜污染的影响[J]. 水处理技术, 2007, 33(10): 7–13. Liu M, Wang Z. EPS extraction, chemical analyses and its effect on membrane fouling[J]. Technology of Water Treatment, 2007, 33(10): 7–13. DOI: 10.3969/j.issn.1000-3770.2007.10.003 |
| [17] | 王兴春, 杨致荣, 王敏, 等. 高通量测序技术及其应用[J]. 中国生物工程杂志, 2012, 32(1): 109–114. Wang X C, Yang Z R, Wang M, et al. High-throughput sequencing technology and its application[J]. China Biotechnology, 2012, 32(1): 109–114. |
| [18] | 周娜. 环境交替下污泥膨胀机理与控制研究[D]. 西安: 西安建筑科技大学, 2009. Zhou N. Study on mechanisms and control strategies of sludge bulking in alternating anaerobic-aerobic-anoxic environment[D]. Xi'an:Xi'an University of Architecture and Technology, 2009. |
| [19] | Costerton J W, Irvin R T, Cheng K J. The bacterial glycocalyx in nature and disease[J]. Annual Review of Microbiology, 1981, 35(1): 299–324. DOI: 10.1146/annurev.mi.35.100181.001503 |
| [20] | Wang X H, Zhang H M, Yang F L, et al. Long-term storage and subsequent reactivation of aerobic granules[J]. Bioresource Technology, 2008, 99(17): 8304–8309. DOI: 10.1016/j.biortech.2008.03.024 |
| [21] | McSwain B S, Irvine R L, Hausner M, et al. Composition and distribution of extracellular polymeric substances in aerobic flocs and granular sludge[J]. Applied and Environmental Microbiology, 2005, 71(2): 1051–1057. DOI: 10.1128/AEM.71.2.1051-1057.2005 |
| [22] | Miksch K, Kończak B. Distribution of extracellular polymeric substances and their role in aerobic granule formation[J]. Chemical and Process Engineering, 2012, 33(4): 679–688. |
| [23] | Zhu L, Zhou J, Lv M, et al. Specific component comparison of extracellular polymeric substances (EPS) in flocs and granular sludge using EEM and SDS-PAGE[J]. Chemosphere, 2014, 121(5): 26–32. |
| [24] | 常青. 活性污泥中胞外聚合物的组成及功能分析[D]. 西安: 西安建筑科技大学, 2014. Chang Q. Composition and function of extracellular polymeric substances in the activated sludge[D]. Xi'an:Xi'an University of Architecture and Technology, 2014. |
| [25] | 包常华. 污水生物处理过程中胞外聚合物的生成与控制[D]. 济南: 山东建筑大学, 2007. Bao C H. Production and controlling of extracellular polymeric substances in wastewater treatment process[D]. Ji'nan:Shandong Jianzhu University, 2007. |
| [26] | 曹秀芹, 赵自玲. 胞外聚合物(EPS)构成的影响因素分析[J]. 环境科学与技术, 2010, 33(S2): 420–424. Cao X Q, Zhao Z L. Analysis of influencing factor on the component of extracellular polymeric substance (EPS)[J]. Environmental Science & Technology, 2010, 33(S2): 420–424. |
| [27] | 侯红娟, 王洪洋, 周琪. 进水COD浓度及C/N值对脱氮效果的影响[J]. 中国给水排水, 2005, 21(12): 19–23. Hou H J, Wang H Y, Zhou Q. Effect of influent COD concentration and C/N ratio on denitrification[J]. China Water & Wastewater, 2005, 21(12): 19–23. DOI: 10.3321/j.issn:1000-4602.2005.12.005 |
| [28] | 金春姬, 佘宗莲, 高京淑, 等. 低C/N比污水生物脱氮所需外加碳源量的确定[J]. 环境科学研究, 2003, 16(5): 37–40. Jin C J, She Z L, Ko K S, et al. Determination of demand of external carbon source for biological nitrogen removal of wastewater with low C/N ratio[J]. Research of Environmental Sciences, 2003, 16(5): 37–40. |
| [29] | 田建强. 反硝化过程中亚硝酸盐积累的影响因素[J]. 有色冶金设计与研究, 2008, 29(3): 42–44. Tian J Q. Influencing factors of nitrite accumulation during denitrification[J]. Nonferrous Metals Engineering & Research, 2008, 29(3): 42–44. |
| [30] | 曹相生, 钱栋, 孟雪征. 乙酸钠为碳源时的污水反硝化规律研究[J]. 中国给水排水, 2011, 27(21): 76–79. Cao X S, Qian D, Meng X Z. Characteristics of denitrification process with sodium acetate as sole carbon source[J]. China Water & Wastewater, 2011, 27(21): 76–79. |
| [31] | Oh J, Silverstein J A. Acetate limitation and nitrite accumulation during denitrification[J]. Journal of Environmental Engineering, 1999, 125(3): 234–243. DOI: 10.1061/(ASCE)0733-9372(1999)125:3(234) |
| [32] | 高景峰, 彭永臻, 王淑莹, 等. 不同碳源及投量对SBR法反硝化速率的影响[J]. 给水排水, 2001, 27(5): 55–58. Gao J F, Peng Y Z, Wang S Y, et al. Effects of carbon sources and dosages on the denitrification rate in SBR process[J]. Water & Wastewater Engineering, 2001, 27(5): 55–58. |
| [33] | Kapley A, Prasad S, Purohit H J. Changes in microbial diversity in fed-batch reactor operation with wastewater containing nitroaromatic residues[J]. Bioresource Technology, 2007, 98(13): 2479–2484. DOI: 10.1016/j.biortech.2006.09.012 |
| [34] | 王硕. 低温好氧颗粒污泥形成过程及其特性研究[D]. 哈尔滨: 哈尔滨工业大学, 2013. Wang S. The study of aerobic garnular sludge on formation process and characteristics at low temperature[D]. Harbin:Harbin Institute of Technology, 2013. |
| [35] | Tan Z, Reinhold-Hurek B. Dechlorosoma suillum Achenbach, et al. 2001 is a later subjective synonym of Azospira oryzae Reinhold-Hurek and Hurek 2000[J]. International Journal of Systematic and Evolutionary Microbiology, 2003, 53(Pt 4): 1139–1142. |
| [36] | 洪武林, 谢学辉, 于文娟, 等. 假单胞菌P1-1脱氮除磷特性及其动力学研究[J]. 环境科学与技术, 2012, 35(10): 29–33. Hong W L, Xie X H, Yu W J, et al. Nitrogen and phosphorus removal characteristics and kinetics of pseudomonas sp. strain p1-1[J]. Environmental Science & Technology, 2012, 35(10): 29–33. DOI: 10.3969/j.issn.1003-6504.2012.10.007 |
| [37] | 魏清娟. 低温脱氮菌剂的制备及其强化人工湿地脱氮效能研究[D]. 哈尔滨: 哈尔滨工业大学, 2015. Wei Q J. Preparation of low temperature nitrogen removal agent and its bioaugmentation efficiency in the constructed wetland[D]. Harbin:Harbin Institute of Technology, 2015. |