 2017, Vol. 38
2017, Vol. 38

2. 中国科学院亚热带农业生态研究所, 亚热带农业生态过程重点实验室, 中国科学院桃源农业生态试验站, 长沙 410125;
3. 中国科学院大学, 北京 100049
 , ZHANG Miao-miao1 , QIN Hong-ling2 , CHEN Chun-lan2 , WANG Juan1 , WEI Wen-xue2 , LI Yong1
, ZHANG Miao-miao1 , QIN Hong-ling2 , CHEN Chun-lan2 , WANG Juan1 , WEI Wen-xue2 , LI Yong1
2. Key Laboratory of Agro-ecological Processes in Subtropical Regions, Taoyuan Station of Agro-ecology Research, Institute of Subtropical Agriculture, Chinese Academy of Sciences, Changsha 410125, China;
3. University of Chinese Academy of Sciences, Beijing 100049, China
N2O被联合国气候变化框架公约列为仅次于CO2和CH4的管制温室气体,其全球增温潜势 (GWP) 是CO2的298~310倍,其中农业是N2O排放源之一,约占全球每年地球表面总N2O排放的20%~30%,而占人为活动中排放N2O的60%,且仍在继续增加[1].而农业土壤N2O的产生主要是硝化作用与反硝化作用的结果.
影响农田土壤N2O排放的主要因素包括土壤温度、土壤水分、土壤质地、土壤pH及施肥与耕作制度等[2],目前有许多关于N2O排放的研究,但大多数研究都集中在生长季节,而冬季N2O排放容易被忽略,主要是因为冬季温度较低同时降雨量较少,但据徐文彬等[3]的估计,北半球热带-亚热带地区冬闲农田是一个重要的尚未确定的N2O排放源,粗略估计N2O (以N2O-N计) 年释放量为0.75Tg;而旱地土壤冬季往往种植农作物,例如, 油菜、小麦等作物,伴随有施肥过程的产生,都会导致有高N2O排放的可能.另外,冬季的温度虽然较低,但Holtan-Hartwig等[4]研究却表明,即使在低温下也可能存在高N2O排放.伍延正等[5]在湖南桃源已有的研究表明,冬闲稻田和油菜地在冬季N2O排放占全年的贡献较大,因此有必要采取合理的措施减少冬闲稻田和冬季油菜地N2O排放.
间、施用有机肥、推广长效缓释肥料、施用硝化抑制剂及精准农业等方式都能减少N2O排放[6].施用硝化抑制剂被认为是最有效的N2O减排措施之一.已报道的硝化抑制剂有双氰胺 (DCD)、2-氯-6-(三氯甲基) 吡啶 (nitrapyrin)、3, 4二甲基吡唑磷酸盐 (DMPP)、脒基硫脲 (ASU)、2-胺-4-氯-6甲基嘧啶 (AM)、2-巯基苯并噻唑 (MBT) 和乙炔等.其中DCD是较为常用的一类硝化抑制剂.主要作用机制是抑制硝化反应的第一步,抑制NH4+氧化为NO3-,而N2O可由这一步产生,在减少NO3-产生的同时也能抑制反硝化过程中N2O的产生[7],所以能够减少N2O的排放.关于DCD减少N2O排放已有诸多报道.周礼恺等[8]的研究表明硝化抑制剂DCD和脲酶抑制剂HQ联合施用能有效减少N2O在水稻生长期的排放,与对照组相比,在施加DCD和HQ后,总N2O的排放减少了1/3. Ma等[9]在水稻田中的研究表明,施加DCD后,年总排放量减少了29.5%~31.3%. Di等[10]的研究表明,在新西兰高氮草地土壤中施加DCD能够有效减少N2O排放.但是关于DCD对冬闲稻田和冬季作物的影响则鲜见报道,冬季温差变化较大,可能存在冻融交替过程等因素也会促进其他过程产生N2O,同时DCD的抑制作用受温度和水分影响较大[11].有研究表明,DCD在高含水量下没有抑制效果[12].与此同时,也有诸多报道关于DCD抑制N2O排放机制的研究,例如Di等[10]在碱性草地土壤中研究表明,DCD抑制了N2O排放主要是抑制了氨氧化细菌 (AOB) 的数量和土壤硝化活性,而对氨氧化古菌 (AOA) 无明显影响;Zhang等[13]在5个强酸性土壤 (pH<4.5) 中施加DCD后明显抑制了AOA的生长和土壤硝化活性,而对AOB无显著影响.但上述关于酸性土壤的研究主要集中于室内培养实验,且未添加外源氮肥.有研究表明,在高氮施肥条件下,酸性蔬菜土壤的氨氧化过程中,AOB比AOA作用更大[14].因此研究DCD在冬闲稻田和冬季油菜地N2O排放的影响能够为更全面评估DCD的抑制效果及抑制机制提供理论支持.
本研究于中科院桃源农业试验站,设置冬闲稻田和油菜地这2个实验小区,分别设置施加DCD处理,检测DCD对冬闲稻田和油菜地N2O排放的影响以及相应的AOA和AOB群落组成和丰度的响应.
1 材料与方法 1.1 实验区概况土壤样品采自中国科学院桃源农业生态试验站.该实验场地处湖南省桃源县漳江镇宝洞峪村 (111°26′E,28°55′N),位于武陵山区向洞庭湖平原过渡的丘岗地带.年均气温16.5℃,年降雨量1 440 mm (图 1),日照1 520 h,土壤类型为第四纪红土发育的红壤.本实验设在2个不同土地利用类型实验区:冬闲稻田、油菜地.冬闲稻田土壤常年种植的是双季稻,油菜地土壤夏季种植玉米, 冬季则种植油菜.

|
图 1 采样期间气温和降水量变化 Fig. 1 Dynamics of precipitation and air temperature during the sampling period |
气体的采集采用静态箱法,采样箱有箱盖和底座两个部分,箱盖是由PVC材料做成为圆柱体,尺寸为直径50 cm,高度为15 cm.根据作物生长可适时增加底座 (底座和箱盖都是圆形,箱盖的高度一定,通过在底座上再扣一个底座,在底座外延加水密封,达到增加整体高度的目的),箱盖外包有保温层,采样时箱内温度保持与箱外温度大体一致.箱顶上端安装有1个空气搅拌风扇和气体样品采集口.在冬闲稻田和油菜地每个处理摆放有4个采气的箱子.采样时,油菜地的采样箱罩住油菜,每个采样箱3株.
箱上安装有橡胶塞,用注射器抽取气体.每隔10 min采集一次,总共收集5次.用12.5 mL的真空瓶收集气体,N2O利用气相色谱分析.采样的时间一般在上午09:00~11:00进行,这一时段的排放通量与日平均通量相当并且操作性强.采样时将箱盖放在底座边缘四周的凹槽中,加水密封,扣箱后用50 mL塑料注射器于0、10、20、30、40 min抽取箱内气体.同时利用水分温度测量仪观测0~15 cm深度的体积含水量和15 cm深度的土壤温度.采样频率为:平常采样为1周一次,遇到强降雨或者下雪天气,一般选择雨雪后第二天加采一次,因为这样的天气后N2O往往有脉冲式排放.其他时间依据季节变化适当调整取样频率.观测期为2010年10月29日~2011年3月19日,这段时间为晚稻收割后至春季翻耕前的一段时间,定义为冬闲期.采样期间的气温和降水量变化见图 1.实验开始的前两天在冬闲稻田和油菜地分别施入DCD,DCD的施入量为10 kg·hm-2,将一定量的DCD溶入水中,喷洒于相应土地利用类型表面.
采完气样后再在每个小区相应地采集3份土壤,每份土壤都是用“S”型法取样.带回实验室分析其NH4+-N和NO3--N.同时在实验开始前和1月1日采集 (实验中期)2次土样混匀后分装,分子实验样品用液氮速冻置于-80℃冰箱保存,理化分析样品置于4℃保存.将1月1日定为实验中期采样主要是因为以前的报道DCD的半衰期是30~49 d左右[15], 考虑到冬闲期温度较低,所以往后推迟了10 d.
1.3 土壤基本理化性质土壤基本理化测定方法如下.
有机质测定:重铬酸钾容量法[16].
硝、铵态氮测定:称取10 g鲜土,加入50 mL 2 mol·L-1 KCl振荡浸提1 h,过滤,采用连续流动分析仪测定 (Flastar 5000 Analyzer)[16].
速效磷测定:0.5 mol·L-1 NaHCO3浸提、钼锑抗比色法测定[16].
速效钾测定:火焰光度法[16].
全钾测定:NaOH熔融,火焰光度法[16].
pH测定:将土壤经风干过筛处理后,称取20 g风干土样于50 mL烧杯内,加入20 mL试剂水盖上表玻璃,并且持续搅拌悬浮液5 min.若含有吸水性的土壤或盐类等其它复杂基质,可额外加入更多试剂水,并记录加入水量.静置悬浮约1 h,使悬浮液的大部分固体沉淀,必要时利用过滤或离心取得水相层,以pH仪电极测定其pH值[16].
全氮测定:半微量开氏法[16].土壤容重测定:环刀法[16].土壤基本理化性质如表 1.
|
|
表 1 土壤基本理化性质 Table 1 Physico-chemical properties of the soil |
1.4 气体样品测定与计算
用气相色谱仪分析N2O的浓度;N2O检测器为ECD (电子捕获检测器),分离柱内填充料为80~100目porpakQ,载气为N2,流量为30 mL·min-1,检测器温度为300℃,分离柱温度为55℃; 通过测定5个气样浓度进行线性回归,得出气体排放速率, 进而求出气体的排放通量[17].
N2O的排放通量计算:

|
式中, F为N2O排放通量 (以N计) [μg·(m2·h)-1];ρ为标准状态下N2O的密度 (1.98 kg·m-3);V为密闭箱内有效的空间体积 (m3);A为密闭箱覆盖的水面面积 (m2);Δc为N2O气体浓度差 (表示为占空气的体积分数);Δt为采样的时间间隔 (h);T为采样时密闭箱的温度 (℃).
N2O累计排放量计算:
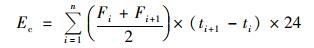
|
式中,Ec为N2O累计排放量 (kg·hm-2);F为N2O排放通量[μg·(m2·h)-1];ti+1-ti为第i和i+1次采样的时间间隔 (d);n为观测期间总测定次数.
1.5 土壤总DNA提取土壤总DNA提取采用SDS-GITC-PEG方法[18].从0.3 g冷冻干燥土壤中提取DNA后,用30 μL灭菌水溶解DNA,然后用0.8%琼脂糖凝胶电泳检测所提取的DNA片段大小,并用NanoDrop核酸蛋白测定仪 (ND-1000) 测定DNA的浓度和质量.
采用表 2所示的引物PCR扩增AOA和AOB的amoA基因,反应体系为:2×power Taq PCR MasterMix (天根,中国)25μL;上下游引物 (10 μmol·L-1) 各1μL;DNA模板约60 ng;加灭菌双蒸水至50μL. PCR反应条件为:95℃预变性5 min;95℃ 30 s, 55℃ 30 s, 72℃ 30 s, 35个循环,最后在72℃下延伸10 min.
|
|
表 2 amoA基因扩增引物 Table 2 Primers used for amoA amplification |
1.6 土壤细菌丰度和群落结构分析
构建amoA基因的标准曲线.首先构建克隆文库, 然后选择阳性克隆子, 从中吸取10 μL菌液于1 mL含氨苄青霉素的LB培养基中, 37℃摇床培养10 h, 提取重组质粒并测定浓度, 进一步换算成单位体积的拷贝数.10倍梯度稀释作为标准曲线的模板, 标准曲线的范围在102~108.拷贝数通过下列公式计算:
拷贝数 (μL)=6.02×1023×质粒浓度 (ng·μL-1)×10-9/(DNA长度×660)
应用实时荧光定量技术测定amoA基因, 每个样品3个重复, 所用引物见表 2.反应体系包括: 5 ng DNA模板, 10 μmol·L-1的上下游引物各0.4μL, 5μL 2×SYBR Premix Ex TaqTM (TaKaRa, 日本), 最后用双蒸水补足至10 μL.采用三步法进行Q-PCR扩增.具体步骤下: 95℃ 30 s; 40个循环包括95℃ 5 s, 68℃ 15 s, 72℃ 25 s, 40个循环; 最后加上熔解曲线过程 (95℃ 15 s, 60℃ 15 s, 95℃ 15 s).
末端限制性片段多态性 (T-RFLP) 按上述同样条件扩增每个DNA样品, 每个样品重复3次, 经1.0%琼脂糖凝胶电泳后, 切取目的条带, 并用琼脂糖凝胶电泳柱式回收纯化试剂盒 (天根, 中国) 进行纯化回收.产物回收后, 对AOA和AOB的amoA基因扩增产物分别用内切酶MSPI (TaKaRa, 日本) 进行酶切反应.反应体系和条件按各内切酶说明书进行.酶切产物送至Sangni公司 (上海, 中国) 进行T-RFLP分析.
1.7 数据分析处理应用SPSS 17.0统计软件对数据进行方差分析 (ANOVA),差异显著性水平通过最小显著差异法 (LSD) 进行检验,用CANOCO软件对AOA和AOB amoA基因T-RFLP结果进行CA分析.
2 结果与分析 2.1 DCD对土壤基本理化性质的影响DCD对土壤理化性质的影响见图 2. DCD对两种土地利用类型的土壤含水量和土壤温度影响较小,冬闲稻田含水量在整个观测期变化较大 (60.0%~90.2%) 而油菜地的含水量基本低于60%土壤孔隙含水量 (WFPS)(45.1%~60.4%),在1月1日~1月8日之间两个土地利用类型之间的含水量较接近.土壤温度在不同土地利用类型之间变化较小,同时也较小受DCD影响.从图 2可以看出,在整个观测期,DCD的施入对NH4+-N累计有一定效果,相比不施加DCD处理,施加DCD使冬闲稻田NH4+-N含量增加了29.1%,而使油菜地NH4+-N含量增加了25.6%.由于DCD主要是抑制硝化过程,DCD的施入对油菜地NO3--N有一定的抑制效果,施加DCD后,总体NO3--N浓度下降了23.8%;冬闲稻田由于本身NO3--N含量较低,DCD的抑制作用不明显.

|
图 2 DCD对不同土地类型环境因子的影响 Fig. 2 Impact of DCD on environmental factors in different land uses |
从图 3中可以看出,冬闲稻田和油菜地N2O排放主要集中在1月15日之前. DCD对不同土地利用类型N2O排放的抑制也主要集中在这段时期,在整个观测期都能减少不同土地类型N2O的排放,从图 3中可以看出冬闲稻田施加DCD后,除了刚开始出现一个排放峰值外,其他时期排放量都较低,抑制效果主要是在前期,冬季不同土地利用方式及施肥处理:冬闲稻田、冬闲稻田+DCD、油菜地、油菜地+DCD的N2O累计排放量分别为0.392、0.223、0.502、0.394 kg·hm-2.整个观测期冬闲稻田和油菜地的N2O排放分别降低了36.7%和23.6%.对于油菜地来讲,DCD的施入也减少了N2O排放,但是在12月29日,油菜地+DCD处理有一个突然增高,总体减排效果要差于冬闲稻田.

|
图 3 DCD对不同土地类型N2O排放的影响 Fig. 3 Impact of DCD on N2O emission in different land uses |
从图 4中可以看出,施加DCD对AOA和AOB的群落组成都有影响,但是不同土地利用类型影响不一. 图 4(a)表明施加DCD后,冬闲稻田AOA群落组成发生了较大变化而油菜地AOA群落组成变化不大.施加DCD后两种土地类型的优势片段T-RF282bp (功能基因片段被酶切后的长度为282bp) 都有所下降,而T-RF166bp (功能基因片段被酶切后的长度为166bp) 在冬闲稻田中增加却在油菜地减少.在油菜地添加DCD处理中,相比如H1(油菜地刚开始时期) 和H2(油菜地中间采样时期),HD2(油菜地施加DCD处理中间采样时期) 中的126bp和134bp有显著增加.对于AOB来说,施加DCD后,冬闲稻田群落组成变化较小而油菜地的群落组成却发生了较大变化.在冬闲稻田中,优势片段T-RF140bp (功能基因片段被酶切后的长度为140bp) 在S2(冬闲水田中间采样时期) 和SD2(冬闲水田施加DCD处理中间采样时期) 中都有所增加,但SD2中的含量要小于S2.在油菜地中,DCD的添加主要抑制了T-RF166bp片段.从图 4(b)也能更进一步看出施加DCD后对AOA和AOB群落结构的不同影响.

|
(a) AOA和AOB群落组成变化; (b) AOA和AOB T-RFLP CA分析; S1、S2、SD2分别代表冬闲水田刚开始、中间采样时期和施加DCD处理的中间采样时期;H1、H2、HD2分别代表油菜地刚开始、中间采样时期和施加DCD处理的中间采样时期 图 4 DCD对不同土地类型amoA基因结构的影响 Fig. 4 Impact of DCD on AOA and AOB amoA gene structure in different land uses |
施加DCD后,对两种土地类型的AOA和AOB丰度都有影响,从图 5中可以看出,AOB丰度在冬闲水田和油菜地中都受到了抑制,分别减少了73.7%和67.0%,在冬闲水田中,施加DCD后基本完全抑制了AOB的生长,相比如实验初期,添加DCD后,冬闲稻田的AOB丰度基本没有增加. AOA的丰度只是在冬闲稻田中减少了59.3%,在油菜地中反而有所上升.除了油菜地添加DCD处理之外,其他处理相比实验初期,AOA丰度都有所下降.

|
1、2、D2分别代表刚开始采样时期、不施DCD处理的中间采样时期、施DCD处理的中间采样时期;SAOA、HAOA、SAOB、HAOB分别代表冬闲水田AOA、油菜地AOA、冬闲水田AOB、油菜地AOB;基因丰度是每克干土的拷贝数 图 5 DCD对不同土地类型AOA和AOB amoA基因丰度的影响 Fig. 5 Impact of DCD on AOA and AOB amoA gene abundance in different land uses |
DCD的作用机制是抑制亚硝化单细菌属的活性,使施入氮源能够较长时间以NH4+-N形态存在,减少NO2--N的积累,进而控制NO3--N的形成[21]. DCD的施入能够减少不同土地利用类型N2O的排放,对冬闲稻田和油菜地N2O累计排放量分别减少了36.7%和23.6%,故本研究说明施用硝化抑制剂DCD是减少冬闲稻田和冬季油菜地N2O排放的一项有效措施.其他研究者也有类似报导,例如Ma等[9]在水稻田中的研究表明,施加DCD后年总排放量减少了29.5%~31.3%, Boeckx等[22]通过室内盆栽小麦和水稻后发现,相对于仅施尿素处理,添加DCD可减少小麦旱作土壤22%的N2O总排放量,还可减少水稻土壤47%的N2O总排放量. Smith等[23]研究表明牧草地施用DCD后N2O年排放总量减少30%~45%,Qiu等[24]研究表明夏季施用DCD后牧草地N2O排放总量减少了40%,冬季则减少了69%.Ruser等[25]综合以前的研究表明,硝化抑制剂的综合减排效率是35%,本研究的结果与这个结果较接近.但也有研究表明DCD在高含水量下无抑制效果[12],虽然冬闲稻田含水量较高,但整个监测期硝态氮含量很低,N2O产生主要来源于硝化过程,所以本研究施入DCD能有效抑制N2O排放.虽说冬闲水田的含水量较高,能加快DCD的降解,但是其抑制N2O排放的时间较长,主要是由于冬闲期温度较低.本研究中油菜地N2O减排效果要小于冬闲稻田添加DCD处理,一个重要的原因是在观测期内,12月29日有一个添加DCD比不添加DCD排放量要高的点.油菜是一个喜硝性作物[26],12月29日温度突然降低到接近零度,减少了油菜对硝态氮的吸收,从而有利于反硝化菌产生N2O,而DCD对反硝化菌无抑制作用[27].与此同时,这期间的降雨也会减弱表层土壤DCD含量从而减弱抑制效果,不添加DCD处理的油菜地铵态氮的含量也相对较高有利于表层土壤硝化过程产生N2O.总的来说,DCD对N2O的抑制作用比较一致,都能减少冬闲稻田和冬季油菜地N2O的排放,但是其抑制效果随着不同土地利用类型以及环境因素的变化而有所不同.
土壤中N2O的产生,主要是由于硝化过程和反硝化过程共同作用的结果,而硝化过程和反硝化的产生依赖于土壤水分含量以及底物含量.不同的研究结果表明,DCD的施入能够抑制N2O的排放,主要是抑制了硝化过程NH4+-N向NO3--N转化,从而抑制了相应的AOA或者AOB生长[25].但关于DCD施入对AOA和AOB的抑制作用报道并不一致. Zhang等[13]发现,中国酸性土壤中氨氧化过程主要由AOA介导,并首次指出DCD可有效降低AOA的数量,而且完全抑制其氧化NH3和固定CO2的活性,对AOB则没有显著影响.杨扬等指出[27],DCD处理可显著减少两种菜地土壤AOB的丰度,而对细菌16S rRNA和反硝化基因nirK的丰度没有显著影响.也有其他的报道表明,施用DCD 7 a的土壤中细菌和古菌16S rRNA基因丰度也与未施用土壤无显著差异[28].从图 4可以看出,水稻田和旱地油菜的AOA和AOB的群落呈现不同的变化趋势,且DCD对他们的抑制作用不一.这个一方面原因是冬闲稻田和油菜地的AOA和AOB的群落结构组成有显著地不同;另外一方面也跟AOA和AOB利用不同来源的NH4+-N有关. AOA利用的NH4+-N主要来源于从有机物矿化出来的氮[29],而AOB主要来源于施加无机肥中的氮[10, 30].在冬闲稻田中,土壤中的无机氮含量较低,主要来源于土壤矿化产生的氮,DCD的施入抑制了NH4+-N向NO3--N转化,由于冬闲稻田较高的含水量以及存在的冻融交替过程,影响土壤的矿化过程,从而对AOA的群落结构造成较大影响[31],而冬闲稻田在整个观测期没有施入外源的无机氮,因而对AOB的影响较小. DCD的施入同时都抑制了AOA和AOB的丰度,张慧等[32]在桃源的实验同时也表明,AOA和AOB积极参与了土壤的硝化过程,说明AOA和AOB都参与了冬闲稻田的硝化过程.
在旱地油菜中,DCD的施入显著改变了AOB的群落结构和丰度,而对AOA的群落结构和丰度影响较小. Zhong等[14]在酸性蔬菜土壤中施入不同含量的氮肥,AOB的群落结构变化比AOA要大,AOB在这些酸性蔬菜土壤的氨氧化过程中起着更为重要的作用.由于在观测期伴随的施肥过程,同时温度和水分的变化以及不同生长阶段对氮的需求的差异也影响油菜对无机氮的吸收[33],导致土壤无机氮变化较大,而土壤矿化氮变化较少,基于AOA和AOB利用不同来源的氮肥,所以导致AOB的群落结构变化比AOA要大,也使AOB丰度更易受DCD的抑制.本研究中油菜地的AOA功能基因丰度在施加DCD一段时间后有所上升.相比于常规施肥处理,添加DCD能够提高油菜的产量[34],AOA丰度的增加可能更多受根际分泌物影响[35].
另外,有研究表明,当土壤的WFPS低于60%时,产生的N2O以硝化作用为主,而反硝化作用产生的N2O随着土壤水分增加而上升;WFPS在60%~80%范围内时,N2O排放作用最强[36].本研究中,冬闲水田的WFPS为60%~90%,较有利于反硝化过程产生N2O.由于冬闲水田本身的NO3--N底物较低,需要硝化过程提供底物,DCD的施入抑制了硝化过程,从以上分析来看,对AOA和AOB都有抑制,但对AOA的抑制效果更强,从而间接抑制了反硝化过程产生N2O[37].油菜地的WFPS为45%~60%左右,N2O产生主要来源于硝化过程,从以上分析来看,DCD的施入主要是抑制了硝化过程中的AOB活性,从而达到减少N2O排放的目的.
4 结论(1) DCD对两种土地类型冬季N2O排放都有一定的抑制作用,对冬闲稻田的抑制效果要大于油菜地.
(2) DCD的施入显著改变了冬闲稻田AOA的群落结构而对AOB的群落结构影响较小.但对AOA和AOB的丰度都有明显的抑制效果,间接抑制了N2O排放.
(3) DCD的施入显著改变了油菜地AOB的群落结构同时明显抑制了AOB的丰度,对N2O的抑制效果主要是抑制了AOB的活性.
| [1] | IPCC. Climate change 2007:changes in atmospheric constituents and in radiative forcing[EB/OL]. http://www.ipcc.ch/pdf/assessment-report/ar4/wg1/ar4-wg1-chapter2.pdf, 2008-01-17. |
| [2] | 谢立勇, 叶丹丹, 张贺, 等. 旱地土壤温室气体排放影响因子及减排增汇措施分析[J]. 中国农业气象, 2011, 32(4): 481–487. Xie L Y, Ye D D, Zhang H, et al. Review of influence factors on greenhouse gases emission from upland soils and relevant adjustment practices[J]. Chinese Journal of Agrometeorology, 2011, 32(4): 481–487. |
| [3] | 徐文彬, 洪业汤. 冬闲水田土壤——一个重要的尚未确定的N2O释放源[J]. 环境科学研究, 1999, 12(3): 42–45. Xu W B, Hong Y T. Paddy fields following a dry season fallow:a potential important N2O source[J]. Research of Environmental Sciences, 1999, 12(3): 42–45. |
| [4] | Holtan-Hartwig L, Dörsch P, Bakken L R, et al. Low temperature control of soil denitrifying communities:kinetics of N2O production and reduction[J]. Soil Biology and Biochemistry, 2002, 34(11): 1797–1806. DOI: 10.1016/S0038-0717(02)00169-4 |
| [5] | 伍延正, 张苗苗, 秦红灵, 等. 不同土地利用方式下冬季N2O排放及其影响因素[J]. 环境科学, 2013, 34(8): 2968–2968. Wu Y Z, Zhang M M, Qin H L, et al. N2O flux in winter and its affecting factors under different land use patterns[J]. Environmental Sciences, 2013, 34(8): 2968–2968. |
| [6] | Hadi A, Jumadi O, Inubushi K, et al. Mitigation options for N2O emission from a corn field in Kalimantan, Indonesia[J]. Soil Science and Plant Nutrition, 2008, 54(4): 644–649. DOI: 10.1111/j.1747-0765.2008.00280.x |
| [7] | Subbarao G V, Ito O, Sahrawat K L, et al. Scope and strategies for regulation of nitrification in agricultural systems-challenges and opportunities[J]. Critical Reviews in Plant Sciences, 2006, 25(4): 303–335. DOI: 10.1080/07352680600794232 |
| [8] | 周礼恺, 徐星凯, 陈利军, 等. 氢醌和双氰胺对种稻土壤N2O和CH4排放的影响[J]. 应用生态学报, 1999, 10(2): 189–192. Zhou L K, Xu X K, Chen L J, et al. Effect of hydroquinone and dicyandiamide on N2O and CH4 emissions from lowland rice soil[J]. Chinese Journal of Applied Ecology, 1999, 10(2): 189–192. |
| [9] | Ma Y C, Sun L Y, Zhang X X, et al. Mitigation of nitrous oxide emissions from paddy soil under conventional and no-till practices using nitrification inhibitors during the winter wheat-growing season[J]. Biology and Fertility of Soils, 2013, 49(6): 627–635. DOI: 10.1007/s00374-012-0753-7 |
| [10] | Di H J, Cameron K C, Shen J P, et al. Nitrification driven by bacteria and not archaea in nitrogen-rich grassland soils[J]. Nature Geoscience, 2009, 2(9): 621–624. DOI: 10.1038/ngeo613 |
| [11] | 孙志梅, 武志杰, 陈利军, 等. 硝化抑制剂的施用效果、影响因素及其评价[J]. 应用生态学报, 2008, 19(7): 1611–1618. Sun Z M, Wu Z J, Chen L J, et al. Application effect, affecting factors, and evaluation of nitrification inhibitor:a review[J]. Chinese Journal of Applied Ecology, 2008, 19(7): 1611–1618. |
| [12] | 白雪, 夏宗伟, 郭彦玲, 等. 硝化抑制剂对不同旱地农田土壤N2O排放的影响[J]. 生态学杂志, 2012, 31(9): 2319–2329. Bai X, Xia Z W, Guo Y L, et al. Effects of nitrification inhibitors on N2O emission from different upland agricultural soils[J]. Chinese Journal of Ecology, 2012, 31(9): 2319–2329. |
| [13] | Zhang L M, Hu H W, Shen J P, et al. Ammonia-oxidizing archaea have more important role than ammonia-oxidizing bacteria in ammonia oxidation of strongly acidic soils[J]. The ISME Journal, 2012, 6(5): 1032–1045. DOI: 10.1038/ismej.2011.168 |
| [14] | Zhong W H, Bian B Y, Gao N, et al. Nitrogen fertilization induced changes in ammonia oxidation are attributable mostly to bacteria rather than archaea in greenhouse-based high N input vegetable soil[J]. Soil Biology and Biochemistry, 2016, 93: 150–159. DOI: 10.1016/j.soilbio.2015.11.003 |
| [15] | Chaves B, Opoku A, De Neve S, et al. Influence of DCD and DMPP on soil N dynamics after incorporation of vegetable crop residues[J]. Biology and Fertility of Soils, 2006, 43(1): 62–68. DOI: 10.1007/s00374-005-0061-6 |
| [16] | 鲍士旦. 土壤农化分析[M]. 北京: 中国农业出版社, 2000: 25-109. |
| [17] | Allen M R, Stott P A, Mitchell J F B, et al. Quantifying the uncertainty in forecasts of anthropogenic climate change[J]. Nature, 2000, 407(6804): 617–620. DOI: 10.1038/35036559 |
| [18] | Chen Z, Luo X Q, Hu R H, et al. Impact of long-term fertilization on the composition of denitrifier communities based on nitrite reductase analyses in a paddy soil[J]. Microbial Ecology, 2010, 60(4): 850–861. DOI: 10.1007/s00248-010-9700-z |
| [19] | Rotthauwe J H, Witzel K P, Liesack W, et al. The ammonia monooxygenase structural gene amoA as a functional marker:molecular fine-scale analysis of natural ammonia-oxidizing populations[J]. Applied and Environmental Microbiology, 1997, 63(12): 4704–4712. |
| [20] | Sahan E, Muyzer G. Diversity and spatio-temporal distribution of ammonia-oxidizing Archaea and Bacteria in sediments of the Westerschelde estuary[J]. FEMS Microbiology Ecology, 2008, 64(2): 175–186. DOI: 10.1111/fem.2008.64.issue-2 |
| [21] | McCarty G W. Modes of action of nitrification inhibitors[J]. Biology and Fertility of Soils, 1999, 29(1): 1–9. DOI: 10.1007/s003740050518 |
| [22] | Boeckx P, Xu X, Van Cleemput O. Mitigation of N2O and CH4 emission from rice and wheat cropping systems using dicyandiamide and hydroquinone[J]. Nutrient Cycling in Agroecosystems, 2005, 72(1): 41–49. DOI: 10.1007/s10705-004-7352-4 |
| [23] | Smith L C, deKlein C A M, Catto W D. Effect of dicyandiamide applied in a granular form on nitrous oxide emissions from a grazed dairy pasture in Southland, New Zealand[J]. New Zealand Journal of Agricultural Research, 2008, 51(4): 387–396. DOI: 10.1080/00288230809510469 |
| [24] | Qiu W H, Di H J, Cameron K C, et al. Nitrous oxide emissions from animal urine as affected by season and a nitrification inhibitor dicyandiamide[J]. Journal of Soils and Sediments, 2010, 10(7): 1229–1235. DOI: 10.1007/s11368-010-0242-2 |
| [25] | Ruser R, Schulz R. The effect of nitrification inhibitors on the nitrous oxide (N2O) release from agricultural soils-a review[J]. Journal of Plant Nutrition and Soil Science, 2015, 178(2): 171–188. DOI: 10.1002/jpln.v178.2 |
| [26] | 田霄鸿, 李生秀. 几种蔬菜对硝态氮、铵态氮的相对吸收能力[J]. 植物营养与肥料学报, 2000, 6(2): 194–201. Tian X H, Li S X. Uptake capacity of several vegetable crops to nitrate and ammonium[J]. Plant Nutrition and Fertilizer Science, 2000, 6(2): 194–201. DOI: 10.11674/zwyf.2000.0211 |
| [27] | 杨扬, 孟德龙, 秦红灵, 等. 硝化抑制剂对蔬菜土硝化和反硝化细菌的影响[J]. 生态学报, 2012, 32(21): 6803–6810. Yang Y, Meng D L, Qin H L, et al. Mechanism of nitrification inhibitor on nitrogen-transformation bacteria in vegetable soil[J]. Acta Ecologica Sinica, 2012, 32(21): 6803–6810. |
| [28] | Guo Y J, Di H J, Cameron K C, et al. Effect of 7-year application of a nitrification inhibitor, dicyandiamide (DCD), on soil microbial biomass, protease and deaminase activities, and the abundance of bacteria and archaea in pasture soils[J]. Journal of Soils and Sediments, 2013, 13(4): 753–759. DOI: 10.1007/s11368-012-0646-2 |
| [29] | Erguder T H, Boon N, Wittebolle L, et al. Environmental factors shaping the ecological niches of ammonia-oxidizing archaea[J]. FEMS Microbiology Reviews, 2009, 33(5): 855–869. DOI: 10.1111/j.1574-6976.2009.00179.x |
| [30] | Zhang L M, Offre P R, He J Z, et al. Autotrophic ammonia oxidation by soil thaumarchaea[J]. Proceedings of the National Academy of Sciences of the United States of American, 2010, 107(40): 17240–17245. DOI: 10.1073/pnas.1004947107 |
| [31] | DeLuca T H, Keeney D R, McCarty G W. Effect of freeze-thaw events on mineralization of soil nitrogen[J]. Biology and Fertility of Soils, 1992, 14(2): 116–120. DOI: 10.1007/BF00336260 |
| [32] | 张慧, 袁红朝, 朱亦君, 等. 不同利用方式对红壤坡地微生物多样性和硝化势的影响[J]. 生态学杂志, 2011, 30(6): 1169–1176. Zhang H, Yuan H Z, Zhu Y J, et al. Microbial diversity and nitrification potential of sloping land red soil under different land use patterns[J]. Chinese Journal of Ecology, 2011, 30(6): 1169–1176. |
| [33] | Malagoli P, Laine P, Rossato L, et al. Dynamics of nitrogen uptake and mobilization in field-grown winter oilseed rape (Brassica napus) from stem extension to harvest:I. Global N flows between vegetative and reproductive tissues in relation to leaf fall and their residual N[J]. Annals of Botany, 2005, 95(5): 853–861. DOI: 10.1093/aob/mci091 |
| [34] | 串丽敏, 赵同科, 安志装, 等. 添加硝化抑制剂双氰胺对油菜生长及品质的影响[J]. 农业环境科学学报, 2010, 29(5): 870–874. Chuan L M, Zhao T K, An Z Z, et al. Effects of adding a nitrification inhibitor dicyandiamide (DCD) on the growth and quality of rape[J]. Journal of Agro-Environment Science, 2010, 29(5): 870–874. |
| [35] | Chen X P, Zhu Y G, Xia Y, et al. Ammonia-oxidizing archaea:important players in paddy rhizosphere soil?[J]. Environmental Microbiology, 2008, 10(8): 1978–1987. DOI: 10.1111/emi.2008.10.issue-8 |
| [36] | Granli T, Bøckman O C. Nitrous oxide from agriculture[J]. Norwegian Journal of Agricultural Sciences, 1994, 12: 127–128. |
| [37] | Li X L, Zhang G B, Xu H, et al. Effect of timing of joint application of hydroquinone and dicyandiamide on nitrous oxide emission from irrigated lowland rice paddy field[J]. Chemosphere, 2009, 75(10): 1417–1422. DOI: 10.1016/j.chemosphere.2009.02.006 |