 2017, Vol. 38
2017, Vol. 38



2. 重庆市三峡库区农业面源污染控制工程技术研究中心, 重庆 400716;
3. 重庆市农业资源与环境研究重点实验室, 重庆 400716
2. Chongqing Engineering Research Center for Agricultural Non-point Source Pollution Control in the Three Gorges Reservoir Area, Chongqing 400716, China;
3. Chongqing Key Laboratory of Agricultural Resources and Environment, Chongqing 400716, China
环境汞 (Hg) 对人体的危害自“水俣病”事件以来逐渐被人们深入认识[1],作为一种“全球性污染物”,Hg广泛分布于大气、土壤以及水体环境中[2].而在水体环境中,汞以甲基汞 (MeHg) 的形式在鱼体富集的现象十分普遍,这使得食用鱼类成为人类Hg暴露的重要途径[2~5].野外同位素示踪实验表明[3]:大气无机汞沉降进入湖泊水域后,仅2个月内就会形成MeHg,并经过食物链进入鱼体.
Hg对鱼体的新陈代谢没有价值,鱼体富集Hg属于被动过程,受到沉积物中Hg甲基化动力学、Hg的食物链转移以及水生生态系统的物理化学特征控制[6].由于水生食物链对MeHg的生物放大作用,鱼体中的Hg含量往往高出其所处水环境许多[7, 8].然而目前对于水环境中Hg形态的生物地球化学过程仍然缺乏掌握[9],这也导致对水环境变化影响鱼体富集Hg的认识不足.
水库作为一种特殊的水环境,由于人为调控水位周期性变化,淹水期洪水携带的泥沙等形成的沉积物,枯水期这些沉积物会落干,致使库区产生大面积的消落带.消落带是库区径流的汇集地带,也成为环境Hg等污染物的汇集区,当淹水发生时消落带会成为水体的Hg污染源[10].三峡水库作为特大型年度调节水库,年度水位落差达30 m,消落带面积约400 km2[11].这些大面积消落带在落干期有可能接受来自地表径流和大气沉降Hg的污染,再次淹水必然会对水体环境和生物产生影响.
许多研究考察了鱼体自身条件对其富集Hg的影响[12~15],主要结论有:体长、体重或年龄是影响鱼体Hg含量种内差异的重要因素,鱼体Hg含量与体长、体重具有正相关性;而食性、营养级水平和体重增长率是影响Hg生物累积种间差异的重要因素,肉食性鱼类或营养级较高的鱼类,体内汞含量通常比较高,而较大的体重增长率意味着生长稀释作用较强,会导致鱼体汞浓度降低.大量研究调查了不同水环境中鱼体的Hg富集情况[6, 12, 15, 16].然而,有关水环境变化对鱼体富集Hg的过程研究比较少见,对于水库这样特殊的水环境更是鲜见报道.同时,由于Hg的甲基化过程主要在水-沉积物界面区域进行[17],因此研究库区消落带沉积物Hg对水生系统和鱼类的影响尤为重要.
本研究以三峡库区重庆段消落带沉积物和江水为材料,进行室内模拟淹水环境实验,考察了不同Hg含量的沉积物,对水体以及鱼体不同部位的影响.
1 材料与方法 1.1 实验材料供试沉积物和水均来自三峡库区重庆段,忠县石宝寨新政村 (N30°25′7.5″, E108°10′5.5″) 消落带.消落带落干时采集沉积物样品和水样带回实验室,沉积物经风干、研磨、过2 mm尼龙筛,最后混匀.沉积物基本性质如表 1所示.取20 kg已过筛的沉积物制备高汞沉积物样品,加入一定量10 mg ·L-1的HgCl2溶液,再加入去离子水调节含水率达饱和状态,搅拌混合均匀后放入1.2 m×0.6 m、内衬为Teflon材料的木缸内,密闭条件下陈化1个月,期间定期搅拌混合2次.陈化后的沉积物经风干,作为高汞沉积物备用,其总汞 (THg) 和甲基汞的含量分别为 (878.74±19.87)μg ·kg-1和1.22 μg ·kg-1.水样基本性质见表 2.
|
|
表 1 供试沉积物的基本理化性质 Table 1 Physicochemical properties of sediments used in this research |
|
|
表 2 供试水样的基本性质 Table 2 Properties of water in the Yangtze River |
供试鱼为斑马鱼,体重 (0.38±0.03) g,体长 (3.90±0.51) cm,实验前用供试水样驯化30 d,期间每2 d喂食一次,饲料购自北京三友创美饲料科技有限公司,THg < 3.57 ng ·kg-1,MeHg < 2.63 ng ·kg-1,喂食量为斑马鱼体重的2%.实验容器由硼硅玻璃制成,尺寸为L60 cm×W 30 cm×H 40 cm.
1.2 实验设计与方法设置3组不同处理,A:对照组,只加入40 L水样;B:原沉积物组,加入6.5 kg原沉积物样品和40 L水样;C:高汞沉积物组,加入6.5 kg高汞沉积物样品和40 L水样.每组处理均设置3个平行.在B组和C组的沉积物上方2 cm处固定一张Teflon网,防止鱼与沉积物直接接触.最后每个处理中各放入60尾斑马鱼.实验温度控制在 (24±1)℃,实验总时长为90 d,期间不给斑马鱼喂食 (根据三峡库区年度水位调节惯例,每年淹水时长90 d的位置对应于海拔172 m左右的消落区,而消落区海拔高度在145~175 m之间,上部消落区由于落干时间更长,沉积物受环境污染的可能性也更大).本实验所使用的玻璃器皿均经过超净处理,用硝酸 (25%,体积分数) 浸泡24 h以上,超纯水清洗后经马弗炉500℃灼烧30 min,冷却后再使用.操作环境保持清洁.
实验期间定期采集鱼样、水样以及沉积物样.以放入斑马鱼的时间点开始计时,于第7、14、28、45、60、75、90 d采集鱼样及沉积物样;于第3 h、12 h、1 d、3 d、7 d、14 d、21 d、28 d、45 d、60 d、75 d、90 d采集水样.对于每个处理,采集鱼样5尾,立即在冰袋上解剖,去除鱼皮,分离头部、内脏和肌肉于自封袋中,放入冷冻真空干燥机干燥,用玛瑙研钵磨成细粉,冷冻保存待测;采用自制的管装采样器在玻璃缸四周及中间多点采集沉积物于自封袋,真空冷冻干燥,研磨并过0.149 mm尼龙筛,冷冻保存待测;采用Teflon管虹吸采集150 mL水样于硼硅玻璃瓶中,加入0.75 mL优级纯浓盐酸酸化,冷藏保存待测.
1.3 样品分析与质量控制对于鱼样及沉积物样品,采用DMA-80固体进样自动测汞仪测定THg含量.按照何天荣等建立的方法[18]测定沉积物样品MeHg,按照阎海鱼等[19]建立的方法测定鱼样MeHg.对于水样,按照USA EPA Method 1631[20]、Method 1630[21]分别测定THg和MeHg.
采用空白实验、平行样品控制及标准物质测定加标回收率进行质量控制.土样标准物质为GBW07405(THg含量:290 ng ·g-1),鱼样标准参考物质为干鱼样NRCC-TORT-2.本实验对于固体样品、水样THg测定加标回收率分别为93%~112%、88%~109%;MeHg测定加标回收率则分别为86%~109%、89%~113%.
2 结果与讨论 2.1 沉积物Hg含量变化实验开始后两组沉积物中的THg开始向水体中迁移,沉积物THg含量逐渐降低,并在第45 d趋于稳定,如图 1(a)所示.实验结束时,原沉积物 (B组) THg含量下降了43.13 ng ·g-1(降幅38.0%),而高汞沉积物 (C组) 则下降了137.61 ng ·g-1(降幅15.7%),C组沉积物THg释放量远高于B组.由于C组THg初始含量很大,虽然其THg含量落差比B组大很多,但其下降幅度却小于B组.通常认为,沉积物间隙水与上覆水中的Hg浓度梯度是Hg在沉积物-水体界面迁移的重要动力[22].本实验在初期时,由于THg浓度梯度的存在,沉积物THg会先进入沉积物间隙水,再沿浓度梯度方向扩散进入上覆水,从而导致沉积物THg含量降低,后期随着浓度梯度逐渐减小,沉积物THg的迁移也减缓,最终达到稳定.

|
图 1 沉积物Hg含量变化 Fig. 1 Variations of Hg levels in sediments |
C组沉积物MeHg的初始含量接近于B组的3倍,见图 1(b),表明C组沉积物在进行陈化时发生了明显的甲基化作用.在实验开始后第7 d,两组沉积物的MeHg含量均明显升高 (独立样本T检验:B组P < 0.05、C组P < 0.10),表明了沉积物中发生了净甲基化作用.这种甲基化作用主要是由于微生物引起的[23],淹水形成的厌氧环境有利于厌氧微生物 (主要为硫酸盐还原菌和铁还原菌) 对Hg的甲基化[24].然而由于第一次采样间隔时间较长,本研究推测实际净甲基化作用发生的时间可能小于7 d. Zhang等[23]曾发现加入外源HgCl2的沉积物,在4℃下经历4 h就会发生明显的微生物甲基化作用.实验7 d之后沉积物MeHg含量基本稳定 (T检验结果:P>0.10),最终B组沉积物MeHg含量升高1.190 ng ·g-1(涨幅320.7%),C组升高0.385 ng ·g-1(涨幅31.5%),B组的增长幅度远大于C组.
值得注意的是,虽然两组沉积物初始THg和MeHg含量差异很大,但实验结束时两组沉积物的MeHg水平并无显著性差异 (P>0.10),实验过程中B组表现出更高的MeHg增长率,这说明沉积物中THg含量并不是MeHg含量的决定性因素,这一点印证了以往的结论[25].已有研究认为:甲基化作用主要依赖于厌氧微生物的新陈代谢[23],同时也需要无机Hg具有生物可利用性,底泥厌氧环境中的HgS沉淀往往会降低生物可利用性[25].在本研究中,受实验条件控制,两组沉积物环境基本一致,因此其微生物数量和活性差异不大,且生物可利用态Hg的水平可能基本一致,最终导致MeHg水平并没有很大的差异.然而沉积物环境的改变有可能会促使微生物活性和数量,以及Hg生物可利用性情况发生变化,导致更多的无机Hg甲基化或者MeHg去甲基化,这需要进一步的实验来验证.
2.2 水体Hg含量变化对照组 (A组) 水体THg浓度基本稳定,两实验组水体THg浓度总体上呈先增后减的趋势,如图 2(a)所示.在实验刚开始的1 d内,两实验组水体THg浓度均异常升高后又回落,这可能是由于淹水初期水体悬浮的沉积物颗粒较多,导致水样颗粒态THg较高所致,随时间推移悬浮颗粒物沉降而相应的颗粒态THg也回到沉积物中,水体THg浓度又降低.沉积物在淹水过程中,水力扰动会导致沉积物释放颗粒态Hg加剧,且沉积物Hg含量越高影响越大,环境风险也越大.在三峡水库水位涨落的过程中,沉积物坡面容易发生侵蚀作用,沉积物颗粒会进入并悬浮于水体[26],这可能导致库区水位上涨时水体颗粒态Hg的升高,高Hg污染的沉积物更易对水体产生这种影响.

|
图 2 水体Hg浓度变化 Fig. 2 Variations of Hg levels in water |
实验开始1 d后,两实验组水体THg浓度开始增加,表明沉积物THg会向上覆水体中扩散,沉积物是上覆水体重要的THg源.值得注意的是:C组的沉积物THg含量约为B组的11倍,但总体来看其上覆水体THg浓度却并未高出B组许多,在实验结束时两组水体THg浓度甚至无显著性差异 (P>0.10),这表明陈化后的高汞沉积物所结合的Hg大部分比较稳定,不易迁移进入水体,反映出沉积物作为水生系统的Hg库对Hg的长期固定作用. Harris等[3]的野外实验也证实了:当外源Hg以大气沉降方式进入湖泊时,大部分最终会稳定地存在于湖底沉积物中.
水体MeHg浓度的动态变化见图 2(b).整个实验周期内,对照组MeHg浓度没有发生显著变化 (P>0.10).对于实验组,第1 d内水体含较多悬浮沉积物颗粒,致使MeHg浓度偏高. 1 d之后,实验组MeHg浓度均呈现先升高再降低的趋势,45 d后MeHg浓度均达到稳定 (P>0.10). MeHg浓度峰值出现在14 d左右,B组达 (0.828±0.153) ng ·L-1,C组达 (1.191±0.256) ng ·L-1,二者无显著性差异 (P>0.10).第28 d时,两实验组水体MeHg浓度均大幅降低,这可能主要与水体中的MeHg光降解作用[27]以及鱼体富集有关.实验结束时B组和C组水体MeHg浓度无显著性差异 (P>0.10).
在第14 d左右时沉积物THg含量达到较高水平,表明此期间Hg从沉积物向水体的扩散作用较强,同时沉积物MeHg含量在第7 d后已达到较高水平的稳定状态,能够作为水体MeHg的源.因此本研究推测水体MeHg浓度的升高主要是由于表层沉积物中的MeHg扩散所致,同时也可以认为实验组沉积物MeHg含量差异不显著导致了上覆水体MeHg浓度差异也不显著.以往研究也认为:水体自身的Hg甲基化作用较弱[28],而表层沉积物具有较强的Hg甲基化能力,同时也是上覆水体重要的MeHg源[29].
2.3 鱼体Hg含量变化食用鱼肉是人类暴露于汞的重要途径,因此笔者将鱼体肌肉作为研究对象.另外,MeHg可通过食物链放大,所以摄食是鱼体暴露于Hg的重要途径,因此考察了作为消化系统的内脏. MeHg具有神经毒性,为了考察Hg通过鱼体血脑屏障进而影响脑部的能力,笔者也将头部纳入研究范围.实验周期中不喂食,鱼体体重增加量微小,不对Hg含量分析构成影响.
图 3展示了各组处理条件下鱼体各部位THg和MeHg含量的动态变化.总体来看,鱼体不同部位THg和MeHg含量的变化规律基本一致.实验结束时,实验组鱼体各部位的THg和MeHg含量均显著升高 (P < 0.01),而对照组除了内脏MeHg没有显著变化 (P>0.10) 以外,其它部位THg和MeHg含量也均有明显升高 (P < 0.05),内脏THg含量有所升高 (P < 0.10).这表明鱼体各部位对Hg均有富集作用,而对照组鱼体Hg含量升高可能是由于其水体本身含有一定量的Hg所致.实验结束时,不同处理条件下,肌肉对THg和MeHg的富集量 (最终含量减去初始含量) 均显著高于头部和内脏 (P < 0.05),而头部和内脏对THg和MeHg的富集量均没有显著性差异 (P>0.10).表明鱼体中肌肉对Hg的富集作用最强.而有关三峡库区鱼体肌肉和内脏的调查也发现了此规律[30].

|
图 3 鱼体各部位Hg含量变化 Fig. 3 Variations of Hg levels in different parts of zebrafish |
本实验结束时,考察肌肉对THg的富集量,呈现C组>B组>A组 (P < 0.01);肌肉对MeHg的富集量也具有同样差异性.内脏对THg的富集量,呈现C组>B组>A组 (A与B、A与C,P < 0.01;B与C,P < 0.05);内脏对MeHg的富集量,有B、C组>A组 (A与B、A与C,P < 0.05;B与C,P>0.10).头部对THg的富集量,有C组>A、B组 (C与A,P < 0.01;C与B,P < 0.05;A与B,P>0.10);而头部对MeHg的富集量,3组之间没有显著性差异.由此可见:含有Hg的沉积物确实会增强鱼体对Hg的富集,并且沉积物Hg含量越高这种加剧鱼体Hg富集的作用越强.但是沉积物Hg含量对鱼体不同部位富集Hg的影响强度不一,具体表现为:对肌肉的影响最为强烈,内脏次之,而对头部最弱.
除了最终结果显示出的差异性外,鱼体各部位富集Hg的过程也有所差异. 图 3(a1) 与3(a2) 显示出,在整个实验周期中,肌肉THg和MeHg含量几乎一直处于增长中;而头部[图 3(c1) 与3(c2)]和内脏[图 3(b1) 与 (b2)]似乎具有先增后减的趋势,且这种趋势在头部中更为明显. B组头部THg含量的最大值 (45 d) 与最终THg含量具有极显著性差异值 (P < 0.01),这确实证明45 d后B组头部THg含量在降低.而这种降低趋势与鱼体不同部位代谢Hg的能力有关,有研究表明:鱼体不同部位代谢Hg的能力呈现腮>脑>内脏>肌肉[31],头部THg含量的显著降低可能主要是由于腮和脑的代谢作用造成的,同时由于肌肉对Hg的代谢能力最弱,所以肌肉中THg和MeHg水平一直在增长.
在整个实验周期中,不考虑各组处理的差异,鱼体3个部位的MeHg和THg含量均呈极显著相关,不同部位MeHg占THg的比例均比较高,并呈现肌肉>内脏>头部,如表 3所示.实际上,各部位THg和MeHg含量高度相似的变化规律 (如图 3) 已经反映出这种强相关性和高MeHg比例.这些现象表明鱼体所富集的Hg形态上主要是MeHg,而对于不同部位,线性方程系数k(表 3) 越大表明富集Hg的过程中,MeHg所占的比例越大.许多调查也发现鱼体肌肉MeHg和THg含量具有极强相关性 (r>0.9),且MeHg比例很高 (45%~100%)[25, 30, 32, 33],这与本研究结果相同.而有关内脏和头部的报道较少甚至没有发现,靳立军等[30]调查三峡库区鱼类时发现鱼体肝、肾、脾中MeHg和THg含量也具有极强相关性,且MeHg比例达40%~50%.
|
|
表 3 各部位MeHg与THg含量相关性分析1) Table 3 Correlation analyses of MeHg and THg concentrations in each part |
生物富集系数 (bioaccumulation factors, BCF) 可以表征鱼体对Hg的富集能力,鱼体对水体Hg的BCF一般用下式来计算[34]:
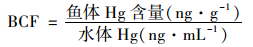
|
以实验结束时鱼体Hg含量和最后两次测定的水体Hg浓度均值为基准,分别计算鱼体各部位对MeHg和无机Hg [鱼体和水体含其它有机Hg形态极少,因此定义无机Hg含量 (浓度)=THg-MeHg]的BCF,结果如表 4所示.
|
|
表 4 生物富集系数对比 Table 4 Comparison of the BCFs |
可见鱼体对MeHg的富集系数总体上要比其他形态的高两个数量级,说明鱼体各部位对MeHg的富集能力都远远超过其他形态的Hg.综上表明,MeHg是导致鱼体富集Hg的直接且最重要原因.不同处理组之间MeHg的富集系数有所差异,但从范围上来看,内脏和头部对MeHg的富集系数在同一个等级,而肌肉则远大于前两者,这表明肌肉是鱼体蓄积Hg的主要部位.本研究的THg富集系数在7.42×103~2.05×104之间,这与一些针对水库鱼类的调查结果[34]相符,没有发现有关MeHg富集系数的调查结果.
有研究发现鱼体中确实存在自身甲基化作用,可将吸收的无机Hg转化为MeHg[31],但是目前普遍认为鱼体内甲基化作用十分微弱可以忽略,鱼体MeHg主要是通过食物链富集以及从水体环境中直接吸收而来[30, 35~37].从水体环境直接吸收的MeHg主要通过鱼鳃进入血液,然后进入其他部位[31].本研究中鱼体Hg应该主要来自于水体吸收,同时水体中的悬浮颗粒物、细小有机体或浮游生物也可能作为鱼的食物,摄食富集可能也占一小部分.
3 结论(1) 消落带沉积物再淹水后,会增加水体THg水平,并在初期致使水体MeHg水平升高,之后又降回初始水平.含Hg沉积物的存在会明显增加鱼体对Hg的富集量.
(2) 当沉积物THg含量更高时,鱼体对THg和MeHg的富集量将显著增加,而水体最终THg和MeHg浓度没有显著增加.
(3) 肌肉对Hg的富集能力最强,而内脏和头部相对较弱且水平相当.鱼体富集的Hg形态主要是MeHg,占THg富集量的比例分别为:肌肉80.1%、内脏79.3%、头部66.7%.
| [1] | Rice K M, Walker E M, Wu M Z, et al. Environmental mercury and its toxic effects[J]. Journal of Preventive Medicine & Public Health, 2014, 47(2): 74–83. |
| [2] | 冯新斌, 陈玖斌, 付学吾, 等. 汞的环境地球化学研究进展[J]. 矿物岩石地球化学通报, 2013, 32(5): 503–530. Feng X B, Cheng J B, Fu X W, et al. Progresses on environmental geochemistry of mercury[J]. Bulletin of Mineralogy, Petrology and Geochemistry, 2013, 32(5): 503–530. |
| [3] | Harris R C, Rudd J W M, Amyot M, et al. Whole-ecosystem study shows rapid fish-mercury response to changes in mercury deposition[J]. Proceedings of the National Academy of Sciences of the United States of America, 2007, 104(42): 16586–16591. DOI: 10.1073/pnas.0704186104 |
| [4] | Bravo A G, Cosio C, Amouroux D, et al. Extremely elevated methyl mercury levels in water, sediment and organisms in a Romanian reservoir affected by release of mercury from a chlor-alkali plant[J]. Water Research, 2014, 49: 391–405. |
| [5] | Driscoll C T, Mason R P, Chan H M, et al. Mercury as a global pollutant:sources, pathways, and effects[J]. Environmental Science & Technology, 2013, 47(10): 4967–4983. |
| [6] | Donald D B, Wissel B, Anas M U M. Species-specific mercury bioaccumulation in a diverse fish community[J]. Environmental Toxicology and Chemistry, 2015, 34(12): 2846–2855. DOI: 10.1002/etc.3130 |
| [7] | Lavoie R A, Jardine T D, Chumchal M M, et al. Biomagnification of mercury in aquatic food webs:a worldwide meta-analysis[J]. Environmental Science & Technology, 2013, 47(23): 13385–13394. |
| [8] | Wang X, Wang W X. Physiologically based pharmacokinetic model for inorganic and methylmercury in a marine fish[J]. Environmental Science & Technology, 2015, 49(16): 10173–10181. |
| [9] | Chételat J, Amyot M, Arp P, et al. Mercury in freshwater ecosystems of the Canadian Arctic:recent advances on its cycling and fate[J]. Science of the Total Environment, 2015, 509-510: 41–66. DOI: 10.1016/j.scitotenv.2014.05.151 |
| [10] | 梁丽, 王永敏, 张成, 等. 三峡水库消落带土壤与优势植物淹水后对土-水系统汞形态的影响[J]. 环境科学, 2016, 37(3): 955–962. Liang L, Wang Y M, Zhang C, et al. Effect of soil and dominant plants on mercury speciation in soil and water system of water-level-fluctuation zone in the three gorges area[J]. Environmental Science, 2016, 37(3): 955–962. |
| [11] | 牛志明, 解明曙. 三峡库区水库消落区水土资源开发利用的前期思考[J]. 科技导报, 1998, 16(4): 61–62, 6. |
| [12] | 朱艾嘉, 许战洲, 柳圭泽, 等. 黄海常见鱼类体内汞含量的种内和种间差异研究[J]. 环境科学, 2014, 35(2): 764–769. Zhu A J, Xu Z Z, Liu G Z, et al. Inner-and inter-species differences of mercury concentration in common fishes from the yellow sea[J]. Environmental Science, 2014, 35(2): 764–769. |
| [13] | Chumchal M M, Hambright K D. Ecological factors regulating mercury contamination of fish from Caddo Lake, Texas, USA[J]. Environmental Toxicology and Chemistry, 2009, 28(5): 962–972. |
| [14] | Power M, Klein G M, Guiguer K R R A, et al. Mercury accumulation in the fish community of a sub-Arctic lake in relation to trophic position and carbon sources[J]. Journal of Applied Ecology, 2002, 39(5): 819–830. DOI: 10.1046/j.1365-2664.2002.00758.x |
| [15] | Kwon S Y, McIntyre P B, Flecker A S, et al. Mercury biomagnification in the food web of a neotropical stream[J]. Science of the Total Environment, 2012, 417-418: 92–97. DOI: 10.1016/j.scitotenv.2011.11.060 |
| [16] | Olejní Dč ková Z, Holešovsky J, Vávrová M, et al. Methylmercury in tissues of the fish from the Svratka river, Czech republic[J]. Fresenius Environmental Bulletin, 2014, 23(12B): 3319–3324. |
| [17] | Ullrich S M, Tanton T W, Abdrashitova S A. Mercury in the aquatic environment:a review of factors affecting methylation[J]. Critical Reviews in Environmental Science and Technology, 2001, 31(3): 241–293. |
| [18] | 何天容, 冯新斌, 戴前进, 等. 萃取-乙基化结合GC-CVAFS法测定沉积物及土壤中的甲基汞[J]. 地球与环境, 2004, 32(2): 83–86. He T R, Feng X B, Dai Q J, et al. Determination of methyl mercury in sediments and soils by GC-CVAFS after aqueous phase ethylation[J]. Earth and Environment, 2004, 32(2): 83–86. |
| [19] | 阎海鱼, 冯新斌, LiangL, 等. GC-CVAFS法测定鱼体内甲基汞的分析方法研究[J]. 分析测试学报, 2005, 24(6): 78–80. Yan H Y, Feng X B, Liang L, et al. Determination of methyl mercury in fish using GC-CVAFS[J]. Journal of Instrumental Analysis, 2005, 24(6): 78–80. |
| [20] | EPA-821-R-02-019 Method 1631, Revision E:mercury in water by oxidation, purge and trap, and cold vapor atomic fluorescence spectrometry[S]. |
| [21] | EPA-821-R-01-020 Method 1630:methyl mercury in water by distillation, aqueous ethylation, purge and trap, and CVAFS[S]. |
| [22] | 贺春凤.三峡库区消落带土壤淹水过程汞释放与甲基化特征的模拟研究[D].重庆:西南大学, 2013. |
| [23] | Zhang L, Planas D. Biotic and abiotic mercury methylation and demethylation in sediments[J]. Bulletin of Environmental Contamination and Toxicology, 1994, 52(5): 691–698. |
| [24] | Barkay T, Wagner-Döbler I. Microbial transformations of mercury:potentials, challenges, and achievements in controlling mercury toxicity in the environment[J]. Advances in Applied Microbiology, 2005, 57: 1–52. DOI: 10.1016/S0065-2164(05)57001-1 |
| [25] | Kannan K, Smith R G Jr, Lee R F, et al. Distribution of total mercury and methyl mercury in water, sediment, and fish from south Florida estuaries[J]. Archives of Environmental Contamination and Toxicology, 1998, 34(2): 109–118. DOI: 10.1007/s002449900294 |
| [26] | 李萍.三峡库区典型消落带水体中汞的时空分布[D].重庆:西南大学, 2013. |
| [27] | 孙荣国, 毛雯, 马明, 等. 水体中甲基汞光化学降解特征研究[J]. 环境科学, 2012, 33(12): 4329–4334. Sun R G, Mao W, Ma M, et al. Characteristics of monomethylmercury photodegradation in water body[J]. Environmental Science, 2012, 33(12): 4329–4334. |
| [28] | 白薇扬.阿哈水库中不同形态汞迁移转化规律的初步探讨[D].贵阳:中国科学院研究生院 (地球化学研究所), 2006. |
| [29] | 何天容.贵州红枫湖汞的生物地球化学循环[D].贵阳:中国科学院研究生院 (地球化学研究所), 2007. |
| [30] | 靳立军, 徐小清. 三峡库区地表水和鱼体中甲基汞含量的分布特征[J]. 长江流域资源与环境, 1997, 6(4): 324–328. Jin L J, Xu X Q. Methylmercury distribution in surface water and fish in the Three-Gorge Reservoir area[J]. Resources and Environment in the Yangtze Valley, 1997, 6(4): 324–328. |
| [31] | 蔺玉华, 张冰艳, 卢健民, 等. 鱼体汞的甲基化及其甲基汞的吸收与代谢[J]. 水产学报, 1994, 18(4): 326–329. Lin Y H, Zhang B Y, Lu J M, et al. Methylation of mercury and methylmercury metabolism in fish body[J]. Journal of Fisheries of China, 1994, 18(4): 326–329. |
| [32] | Grieb T M, Bowie G L, Driscoll C T, et al. Factors affecting mercury accumulation in fish in the upper michigan peninsula[J]. Environmental Toxicology and Chemistry, 1990, 9(7): 919–930. DOI: 10.1002/etc.v9:7 |
| [33] | Kim J P. Methylmercury in rainbow trout (Oncorhynchus mykiss) from Lakes Okareka, Okaro, Rotomahana, Rotorua and Tarawera, North Island, New Zealand[J]. Science of the Total Environment, 1995, 164(3): 209–219. DOI: 10.1016/0048-9697(95)04472-D |
| [34] | 程柳, 毛宇翔, 麻冰涓, 等. 汞在小浪底水库的赋存形态及其时空变化[J]. 环境科学, 2015, 36(1): 121–129. Cheng L, Mao Y X, Ma B J, et al. Speciation and spatial-temporal variation of mercury in the Xiaolangdi Reservoir[J]. Environmental Science, 2015, 36(1): 121–129. |
| [35] | Wiener J G, Cope W G, Rada R G. Mercury accumulation in yellow perch in wisconsin seepage lakes:relation to lake characteristics[J]. Environmental Toxicology and Chemistry, 1990, 9(7): 931–940. DOI: 10.1002/etc.v9:7 |
| [36] | Watras C J, Back R C, Halvorsen S, et al. Bioaccumulation of mercury in pelagic freshwater food webs[J]. Science of the Total Environment, 1998, 219(2-3): 183–208. DOI: 10.1016/S0048-9697(98)00228-9 |
| [37] | 于常荣, 张美昭. 鱼体甲基汞来源及半衰期的研究[J]. 水产科技, 1994(2): 36–38. |