 2017, Vol. 38
2017, Vol. 38

2. 湖北工业大学土木建筑与环境学院, 武汉 430068;
3. 河湖生态修复与藻类利用湖北省重点实验室, 武汉 430068
 , LIU De-fu2,3 , YANG Zheng-jian2,3
, LIU De-fu2,3 , YANG Zheng-jian2,3 , ZHANG Jia-lei2,3 , XU Ya-qian2 , LIU Jin-gao2 , YAN Guang-han1
, ZHANG Jia-lei2,3 , XU Ya-qian2 , LIU Jin-gao2 , YAN Guang-han1
2. College of Civil and Environmental Engineering, Hubei University of Technology, Wuhan 430068, China;
3. Key Laboratory of Ecological Remediation of Lakes and Rivers and Algal Utilization of Hubei Province, Wuhan 430068, China
溶解氧是表征水体污染程度的重要指标[1~3],是维系良好的湖泊水质和健康的最重要条件之一,缺氧以及厌氧条件会对湖泊生态系统产生许多不利影响[4].一般清洁水体溶解氧趋于饱和,当水体受到污染,则溶解氧浓度会大幅降低[5].但同时,浮游植物的光合作用会消耗二氧化碳,释放氧气,使水体中溶解氧增加;若水中溶解氧较低亦会抑制浮游植物的生长[6].有研究发现,大型浅水湖泊太湖溶解氧垂向分布主要受浮游植物、水温层化以及季节影响,垂向分层不稳定[7].深水水库三峡库区其溶解氧表层主要来自浮游植物的光合作用,底层则主要受水动力条件影响[8],呈现表层溶解氧区域饱和,底层出现缺氧状态.有研究表明水体中溶解氧与温度、水体分层、pH、悬浮物、高锰酸盐指数等有重要关系[6, 9~12].而千岛湖属于大型人工深水湖泊,研究其溶解氧与浮游植物分布对水库环境保护具有重要意义.
千岛湖为1959年新安江水电站大坝建成蓄水后所形成的特大山谷型水库,为长三角地区重要水源地,水质常年为Ⅱ类水体[13].近年来,随着经济不断发展,千岛湖局部区域暴发大面积的季节性蓝藻水华[14, 15],引起社会广泛关注.目前,国内对溶解氧与浮游植物的研究大部分集中于三峡库区、浅水湖泊或近海河口区域[16~18].针对千岛湖已有的研究较多关注的是某一特定水深的分布规律.对溶解氧与浮游植物垂向分布规律探讨不多,以及其环境影响因子研究也甚少.因此本文主要研究千岛湖溶解氧与浮游植物垂向分布特征,着重探讨引起溶解氧垂向分布与低氧区形成的原因,通过掌握造成溶解氧与浮游植物特殊分布的原因,以期为后期千岛湖水库环境管理与水资源利用提供科学依据.
1 材料与方法 1.1 采样点设置千岛湖 (又名新安江水库),位于浙江省西部与安徽省南部交界的淳安县境内 (29°22′~29°50′N,118°34′~119°15′E),1959年新安江水电站大坝建成蓄水后所形成的特大山谷型水库,大坝以上控制流域面积为10 442 km2,其中60%位于安徽省境内,40%位于浙江省境内,平均水深34 m,年平均气温17℃[19].如图 1所示于2015年9月,在千岛湖大坝前湖泊区 (29°29′8″N~29°29′18″N,119°12′46″E~119°8′29″E) 间隔4 km左右设置1个采样点,总计5各采样点,分别为S1(XAJ01)、S2(XAJ02)、S3(XAJ03)、S4(XAJ04)、S5(XAJ05).

|
图 1 千岛湖地形图及采样点分布示意 Fig. 1 Topographic map of Thousand-island Lake and distribution of sampling points |
水温 (WT)、pH值 (pH)、浊度 (turbidity)、溶解氧 (DO)、电导率 (conductivity)、水深 (depth)、透明度 (SD)、叶绿素a浓度 (Chl-a)、水下光合有效辐射 (PAR)、水下消光系数 (Ksd)、总氮 (TN)、氨氮 (NH4+-N)、硝氮 (NO3--N)、总磷 (TP)、正磷酸盐 (PO43--P)、可溶性硅酸盐 (D-Si)、碱度、总硬度 (Mg2+、Ca2+)、高锰酸盐指数等.
1.3 实验分析于各监测点表层、中层、底层处 (S1、S2、S3这3个监测点表层0.5 m,中层30 m,底层55 m),用5 L柱状采水器分别采集水样;理化项目分析48 h内在实验室完成,根据文献[20]测定.水温、pH、浊度、溶解氧、电导率、水深等参数由YSI-EXO便携式/在线多参数水质监测仪 (美国) 现场测定,水下光强用International Light 4100光照计 (美国) 测定;透明度用赛氏盘法现场直接测定.浮游植物定量样品,各层取1 L水样,现场用鲁哥氏剂固定,带回实验室沉淀浓缩至50 mL,用显微镜分类计数;浮游植物物种鉴定和计数等参考文献[21].
1.4 数据分析数据采用Excel分析,显著性差异采用SPSS 17.0进行分析.混合层深度 (m) 取与表层水温温差ΔT=0.5℃时所对应的水深[22],采用1%表面光强所对应的深度 (m) 作为真光层深度[23, 24].
2 结果与分析 2.1 溶解氧垂向分布分布特征千岛湖湖泊区溶解氧在不同水深浓度差异较大,呈现明显分层现象 (图 2);表层溶解氧>底层>中层. S3点表层溶解氧略大于其他断面,为8.25 mg ·L-1;其他各点表层DO为 (7.37±0.1) mg ·L-1.垂向上各监测点分布规律一致,0~12 m内溶解氧浓度较高且垂向上无明显差异. 12 m之后出现明显溶解氧突变减小,变化深度达43 m,最大梯度为0.53 mg ·m-1. 14~25 m水深呈现缺氧状态 (DO浓度小于4.00 mg ·L-1[25]),最小值出现在17 m处,溶解氧为1.95 mg ·L-1.至17 m开始溶解氧突变开始递增,平均递增梯度为0.21 mg ·m-1.在40 m水深处出现第3次突变,溶解氧重新沿水深呈现递减.

|
图 2 千岛湖湖泊区溶解氧垂向分布 Fig. 2 Vertical distribution of dissolved oxygen in Thousand-island Lake |
2015年千岛湖坝前湖泊区水温与pH存在显著分层现象 (图 3).水温、pH在湖泊区各监测点垂向分布均呈现“上高下低”现象[图 3(a)和3(b)].水温在表层0~12 m水深无明显变化,该层水体垂向完全掺混,属于混合层水体 (亦为同温层). 12 m之后出现突变,水温开始沿水深方向出现递减;表、底温差为14.3℃,温度梯度为0.55℃ ·m-1,水体出现深度达28 m的“温跃层”. 40 m水深之后水温重新稳定,水温分层消失.湖泊区水体表层呈现微碱性[图 3(b)],中层呈弱酸性,底层则为中性. pH沿水深方向出现明显分层,0~12 m受水体掺混影响,该层水体内pH值无明显差异. 12 m处出现突变减小,最小值在20 m处,为pH 6.70,20 m之后开始回升.各监测点浊度存在空间差异[图 3(c)],坝前区浊度小于其他区域,S4点浊度最大. S1、S2、S3三点在垂向上沿水深呈现先减小后增大趋势,在pH减小区域浊度变小,在pH增大区域内浊度增加,浊度最大相差10 NTU;S4、S5垂向变化较小.湖泊区各测点电导率垂向分布如图 3(d),各监测点表层差异较小,为 (117±2.5) μS ·cm-1;坝前区域稍大于其他断面.电导率垂向上存在明显差异;S1~S3分布规律相似,表层小于底层,最大S1表底相差15 μS ·cm-1.电导率在0~12 m内无明显变化;在12 m之后出现突变呈现沿水深递减,最小值出现在24 m为111.30 μS ·cm-1;24 m之后开始沿水深逐渐增加. S4与S5断面电导率垂向变化稍小,在0~12 m内无明显变化. S4断面在12 m之后至底层沿水深逐渐递减,S5断面在12~20 m出现先减小后增大的变化,最终表底无差异.

|
图 3 千岛湖湖泊区理化因子垂向分布 Fig. 3 Vertical distribution of physical and chemical factors in Thousand-island Lake |
湖泊区各点浮游植物总密度与叶绿素a垂向分布如图 4,水柱内浮游植物表、中、底平均密度分别为156.10×104、262.30×104、161.90×104 cell ·L-1,密度范围在72.50×104~377.30×104 cell ·L-1之间.水柱内浮游植物分层明显,S1、S2、S3这3个点[图 4(a)、4(b)和4(c)]呈现中层区域密度>表层区域>底层区域,S4、S5[图 4(d)和4(e)]则是底层>表层>中层;可以发现浮游植物密度最大区域均在20~30 m水深位置.各点水柱内叶绿素a分布完全受浮游植物密度影响,垂向分布与总密度规律一致.

|
图 4 千岛湖湖泊区浮游植物总密度及叶绿素a垂向分布 Fig. 4 Vertical distribution of phytoplankton density and chlorophyll a in Thousand-island Lake |
千岛湖湖泊区共鉴定出5门34属.各藻门所占比例在不同监测断面垂向分布上差异较小,以硅藻门 (Bacillariophyceae) 为主[如图 5(a)];总计检测出13属,在表、中、底各层所占比重为56%.绿藻门 (Chlorophyta)13属次之,约占总数的30%;蓝藻门 (Cyanophyta)6属,约占总数的10%;裸藻 (Euglena) 与隐藻 (Cryptophyta) 合计占4%左右.监测区水柱内不同水深藻属生物量无明显差异[图 5(b)],主要优势藻属有卵形藻 (Cocconeis)、脆杆藻 (Fragilaria)、色球藻 (Chroococcus)、直链藻 (Melosira)、小环藻 (Cyclotella)、衣藻 (Chlamydomonas),占总数的70%~80%.表层脆杆藻 (Fragilaria)、卵形藻 (Cocconeis) 生物量小于中、底层,衣藻 (Chlamydomonas)、小环藻 (Cyclotella) 则大于中、底层,而色球藻 (Chroococcus)、直链藻 (Melosira) 在水柱内各层分布较均匀.温度可能是影响不同藻种在不同水深分布的重要因素[26],表层水温在25℃左右,适宜以小环藻、衣藻为主的绿藻生长[27].硅藻适宜在13~25℃水温中生长[28],因此脆杆藻,卵形藻在中层水深偏冷环境中生长较好[29].

|
图 5 千岛湖湖泊区浮游植物群落垂向分布及主要优势藻属 Fig. 5 Vertical distribution of phytoplankton community and the main dominant algae in Thousand-island Lake |
千岛湖湖泊区浮游植物垂向分布与各理化因子相关性分析如表 1.浮游植物生物量与DO、pH呈负相关,相关系数分别为0.554、0.515(P<0.001).由于湖泊区各监测点混合层很大,浮游植物在0~20 m内光合作用无法抵消呼吸作用,致使水体内溶解氧降低;呼吸作用释放大量CO2使得水体pH减小.在20~38 m内浮游生物数量减小,水体中的溶解氧与pH逐渐恢复到正常水平. 38 m之后溶解氧与pH主要受水深影响而减小.水温是影响藻类生长的重要因素之一[30, 31],但湖泊区水柱内以硅藻为主要优势藻种,垂向水温在12~25℃,非常适宜硅藻的生长,因此水柱内水温对藻类生物量在垂向分布无明显影响,相关性也不明显.浮游植物生物量与浊度相关关系也不显著.
|
|
表 1 浮游植物生物量与溶解氧、水温、pH、浊度、叶绿素a的相关关系1) Table 1 Correlation of phytoplankton biomass with dissolved oxygen, temperature, pH, turbidity and chlorophyll a |
3 讨论 3.1 千岛湖湖泊区低氧成因分析
湖库中表层水体溶解氧主要来源于空气中氧气的溶解及浮游植物的光合作用,而深水区的溶解氧主要是通过水体的垂直对流扩散获得.千岛湖湖泊区内溶解氧垂向存在明显分层现象,表层0~12 m内溶解氧含量趋于饱和;该区域水体属于同温层 (即混合层) 内,水体掺混完全,各物质与浮游生物均匀分布.水体中溶解氧浓度较高,其主要来源于空气中的氧气溶解及浮游植物光合作用.溶解氧在12 m之后出现突变减小,形成将近14 m的低氧分层区.在20~38 m水深区域内水温温跃梯度减小,且该区域内浮游动物等好氧生物生物量减小,溶解氧消耗量减小,水体溶解氧浓度逐渐增加至正常状态. 38 m之后溶解氧因水深增加而自然下降.在12~20 m区域内衰减明显,主要原因如下.
(1) 千岛湖湖泊区溶解氧与水温在温跃层内存在显著相关关系,相关性系数为0.647(P<0.001);水温突变形成的热力分层,是引起溶解氧突变减小的主要物理因子[32~35].在淡水中不同温度对应的溶解氧浓度计算公式为[36]:
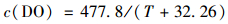
|
(1) |
式中,c(DO) 表示溶解氧浓度;T表示水温.如图 6理论模型所示,在无水温分层影响下,理论溶解氧变化应沿垂向上逐渐递增趋势.而千岛湖湖泊区12~20 m水深区域水温出现突变巨减,温跃深度超过30 m,平均温跃梯度为0.46℃ ·m-1.高梯度的水温分层阻碍了上下水团交换,表层溶解氧无法向下层输移,造成区域内溶解氧不断减小.这与杨丽娜[37]对大辽河溶解氧的研究规律完全吻合. Foley等[38]对英国某湖泊研究与Yin等[39]对珠江口研究均表明温度分层是底层缺氧的主导因素.同时千岛湖湖泊区内高锰酸盐指数平均值为2.42 mg ·L-1,其耗氧作用主要发生在氧化分解. 12~20 m水深区域水温的突变造成区域内富集的还原性物质及营养盐无法向上迁移,这些物质在分解过程中耗氧远超过复氧,区域内水体会逐渐发展为缺氧状态[33, 34]. Boehrer等[40]对世界不同类型湖泊进行分析时发现:“温跃层”出现时水体产生密度差,有机物质与无机物质均会在对应密度的水深积累,这些物质被分解时需要消耗大量的氧.王雨春等[41]对百花湖水库研究发现水体分层限制上下水团在垂直交替过程中耗氧和复氧机制不平衡会引起水体缺氧.

|
图 6 理论溶解氧垂向分布与实际分布对比 Fig. 6 Theoretical vertical distribution and actual distribution of dissolved oxygen |
(2) 千岛湖湖泊区水柱内溶解氧与浮游植物密度相关性系数为0.554(P<0.05),呈现负相关关系.浮游植物生物量直接影响水体中溶解氧的含量[42].杨正健[43]对三峡库区浮游植物的研究中发现浮游植物在真光层内光合作用大于呼吸作用,真光层深度以下时呼吸作用大于光合作用,浮游植物光合作用难以抵消其呼吸作用[44],浮游植物生物量增长受限制.千岛湖湖泊区内真光层平均深度为13.5 m,12~20 m水深内Chl-a平均质量浓度由2.47 μg ·L-1减小至1.08 μg ·L-1,浮游植物生物量衰减明显,该区域水体中营养充分但光照不足,限制了水体的浮游植物生长和初级生产力[35],从而浮游植物呼吸作用强于光合作用,消耗大量O2,释放出CO2.同时从图 7中可以发现12~20 m内浮游植物生物量衰减明显,死亡后的浮游植物在被微生物分解过程中同样需要消耗大量氧气.

|
浮游动物数据文献[45],浮游植物垂向分布特征选用Chl-a为代表 图 7 千岛湖湖泊区溶解氧与相关因子垂向分布模型 Fig. 7 Vertical distribution model diagram of DO and related factors in Thousand-island Lake |
(3) 浮游动物垂向分布规律也是引起溶解氧剧变的一个重要因素. Boehrer等[40]研究发现浮游生物在温跃层为了寻找充足的营养物质而迁移到对应的梯度营养区.如图 7,千岛湖湖泊区浮游动物为获得充足食物在12~20 m内浮游动物大量栖息,浮游动物只进行呼吸作用,消耗大量溶解氧;同时捕食大量的浮游植物,致使浮游植物生物量减小,光合作用产氧也减少.杨丽丽[45]对千岛湖浮游动物研究发现夏季浮游动物分布对溶解氧浓度有重要影响.千岛湖湖泊区白天浮游动物主要分布于10~21 m (97.87%)[46, 47],生物量高达为720个·L-1[48].高密度的浮游动物聚集会消耗水体中大量的氧气,致使监测区水柱内12~20 m水深处溶解氧剧减.
综合分析千岛湖湖泊区溶解氧垂向浓度波动范围在1.95~8.25 mg ·L-1之间,平均浓度为5.10 mg ·L-1.低氧区出现在12~20 m水深,最小值在17 m为1.95 mg ·L-1.通过分析各环境因子可知,如图 7所示,溶解氧垂向变化主要受物理、化学和生物等共同控制,因此可以从水温分层、有机污染物和浮游生物活动等方面综合探讨低氧的形成.表层溶解氧浓度高,主要来自浮游植物光合作用与空气复氧. 12~20 m因溶解氧巨减形成低氧区,分析各监测点 (S1~S5) pH与溶解氧发现,其分布规律在完全一致可知[图 2与图 3(b)],pH的剧变也是引起溶解氧剧变的一个重要因素.水体一般本身具有氧化-还原性质[42, 49],其氧化-还原过程为:O2(气)+4H++4e⇌2H2O (液)[50]. pH值降低,说明H+的活动增大促进水的氧化还原反应向正反应方向进行,则H+与O2反应生成水,使溶解氧降低.监测发现低氧区高锰酸盐指数较高,表明有机物在分解过程中消耗的大量氧气是造成区域内溶解氧衰减过快的重要因素.除此之外低氧区内溶解氧与水温呈现显著正相关关系.该结果表明溶解氧受水温分层影响明显,水温分层决定了垂向水动力交换条件,水温降低会导致温、密跃层的形成,致使上下层水体垂向交换受限制,溶解氧无法由上次输移至下层水体,低氧区水体溶解氧被不断消耗的同时得不到新的补充.低氧区内溶解氧与浮游植物生物量呈现显著相关,该结果表明浮游植物生物量的减少,水体内光合作用产氧减弱.区域内浮游植物呼吸作用大于光合作用,其自身生存需要消耗氧气的同时死亡后残骸的分解过程同样需要消耗大量溶解氧.根据“双峰效应”的滞后性,表层浮游植物充足,浮游动物随之繁盛并随着表层水体掺混至水温分层区聚集,在稳定分层区消耗大量溶解氧,造成低氧区溶解氧进一步衰减.综上所述,水体垂向层化作用等物理过程以及浮游生物的活动直接或间接决定了千岛湖湖泊区低氧区的范围与程度.
3.2 浮游植物垂向分布规律分析千岛湖夏季S1~S3监测点水柱内浮游植物总生物量呈现中层>表层>底层;S4~S5点底层>表层>中层. 0~12 m浮游植物在混合层深度内,表层水体虽然营养 (总氮平均1.41 mg ·L-1;总磷平均0.03 mg ·L-1) 与光照均充足,但是水柱内混合层深度达到11.5 m,混合层内水体垂向紊动,各物质垂向交换频繁,浮游植物无法稳定停留在某一水深内进行光合作用,减弱了其有效光合作用强度,致使净初级生产力较小[51];从而抑制了浮游植物的生长.在12~20 m区域水温分层开始出现,水温突变巨减影响了浮游植物体内酶的活性[31];浮游植物光合作用减弱,从而导致不适宜此低温的暖水性浮游植物生物量[52]在温跃层内剧减 (图 7).在温跃层内热力分层一方面对浮游植物垂向分布形成屏蔽作用,阻碍了其垂向迁移;另一方面造成营养物质垂向交换减弱,形成区域内光照充足而营养物质匮乏的情况[35],浮游植物生物量会因缺少营养物质而大量死亡.同时浮游植物生物量还受到溶解氧的限制影响,Chapman等[53]指出当水体中溶解氧低于4 mg ·L-1时会对浮游生物生长产生抑制作用. 12~20 m内溶解氧突变减小,浮游植物在区域内处于缺氧状态而无法正常生长.浮游动物在12 m水深生物量达到最大也是影响浮游植物在12~20 m内减少的重要因素,浮游动物在该水深内大量捕食,造成溶解氧迅速衰减的同时浮游植物生物量减少明显.
浮游植物生物量最大出现在20~30 m区域内 (S1~S3断面中层,S4~S5底层),主要因为该水深内浮游动物生物量减少,浮游植物被捕食量减少;同时,该区域水体稳定,浮游植物可以稳定接受光照,光合作用效率增加;因此水体中浮游植物生物量明显增加.新安江水电站取水口位于水下25 m左右 (正常高水位108 m,取水口高程73 m[54]),大坝取水时水体横向交换作用加强,较远区域表层部分浮游植物被带入该水深区内 (藻类具有悬浮生长及随流输移的特性[55]).电导率[图 3(d)]在20~30 m区域出现突变,也证明了该区域水体 (20~30 m水深水体) 存在横向水体交换. S1、S2、S3各点底层生物量较小,主要因在深水区,底部缺少光照,浮游植物进行光合作用效率微弱,因此生物量较小.
4 结论(1) 千岛湖湖泊区水温、溶解氧、pH呈现明显分层状态,在0~12 m各指标无明显变化,12 m之后出现温跃层.水柱内水温呈现“表层高底层低”状态,表底温差达为14.3℃.千岛湖湖泊区溶解氧垂向波动范围在1.95~8.25 mg ·L-1之间,平均浓度为5.1 mg ·L-1.低氧区出现在12~20 m水深,最小值在17 m为1.95 mg ·L-1.溶解氧与pH垂向分布规律一致,在温跃层变化较大,出现明显的突变减小.水体垂向层化作用等物理过程以及浮游生物的活动直接或间接决定了千岛湖湖泊区低氧区的范围与程度. pH垂向分布主要受浮游植物在不同水深光合作用与呼吸作用强弱的影响.
(2) 湖泊区内浮游植物共鉴定出5门34属,硅藻门 (Bacillariophyceae) 为主要优势藻种.通过分析发现浮游植物生物量与溶解氧、pH相关性显著.浮游植物垂向上生物量差异明显,S1、S2、S3这3个点浮游植物生物量呈现中层>表层>底层,S4、S5两点浮游植物生物量底层>表层>中层,水柱内浮游植物在20~30 m区域生物量最高.表层主要因水体垂向掺混,在混合层内生长受限制,以及在中层受浮游动物捕食与水体缺氧的影响;在30 m以后主要受光限制影响.
| [1] | 王成善, 胡修棉, 李祥辉. 古海洋溶解氧与缺氧和富氧问题研究[J]. 海洋地质与第四纪地质, 1999, 19(3): 39–47. Wang C S, Hu X M, Li X H. Dissolved oxygen in palaeo-ocean:anoxic events and high-oxic problems[J]. Marine Geology & Qaternary Geology, 1999, 19(3): 39–47. |
| [2] | Neal C, House W A, Jarvie H P, et al. The water quality of the River Dun and the Kennet and Avon Canal[J]. Journal of Hydrology, 2006, 330(1-2): 155–170. DOI: 10.1016/j.jhydrol.2006.04.017 |
| [3] | Engle V D, Summers J K, Macauley J M. Dissolved oxygen conditions in northern Gulf of Mexico estuaries[J]. Environmental Monitoring and Assessment, 1999, 57(1): 1–20. DOI: 10.1023/A:1005980410752 |
| [4] | 张运林. 气候变暖对湖泊热力及溶解氧分层影响研究进展[J]. 水科学进展, 2015, 26(1): 130–139. Zhang Y L. Effect of climate warming on lake thermal and dissolved oxygen stratifications:a review[J]. Advances in Water Science, 2015, 26(1): 130–139. |
| [5] | 方志发, 谢家才. 千岛湖水溶解氧含量与水深影响的探讨[J]. 环境污染与防治, 1999, 21(S1): 65–66. |
| [6] | 张莹莹, 张经, 吴莹, 等. 长江口溶解氧的分布特征及影响因素研究[J]. 环境科学, 2007, 28(8): 1649–1654. Zhang Y Y, Zhang J, Wu Y, et al. Characteristics of dissolved oxygen and its affecting factors in the Yangtze Estuary[J]. Environmental Science, 2007, 28(8): 1649–1654. |
| [7] | 欧阳潇然, 赵巧华, 魏瀛珠. 基于FVCOM的太湖梅梁湾夏季水温、溶解氧模拟及其影响机制初探[J]. 湖泊科学, 2013, 25(4): 478–488. Ouyang X R, Zhao Q H, Wei Y Z. A preliminary exploration of dissolved oxygen based on FVCOM in Meiliang Bay, Lake Taihu and its influence mechanism[J]. Journal of Lake Sciences, 2013, 25(4): 478–488. DOI: 10.18307/2013.0404 |
| [8] | 崔彦萍, 王保栋, 陈求稳. 三峡正常蓄水后长江口叶绿素a和溶解氧变化及其成因[J]. 生态学报, 2014, 34(21): 6309–6316. Cui Y P, Wang B D, Chen Q W. The spatial-temporal dynamics of chlorophyll a and DO in the Yangtze estuary after normal impoundment of the Three Gorges Reservoir[J]. Acta Ecologica Sinica, 2014, 34(21): 6309–6316. |
| [9] | 张运林, 陈伟民, 杨顶田, 等. 天目湖2001~2002年环境调查及富营养化评价[J]. 长江流域资源与环境, 2005, 14(1): 99–103. Zhang Y L, Chen W M, Yang D T, et al. Main physical and chemical factors in Tianmuhu lake, with evaluation of eutrophication from 2001 to 2002[J]. Resources and Environment in the Yangtze Basin, 2005, 14(1): 99–103. |
| [10] | 邹建军, 杨刚, 刘季花, 等. 长江口邻近海域九月溶解氧的分布特征[J]. 海洋科学进展, 2008, 26(1): 65–73. Zou J J, Yang G, Liu J H, et al. Distribution characteristics of dissolved oxygen in the sea area adjacent to the changjiang river estuary in september[J]. Advances in Marine Science, 2008, 26(1): 65–73. |
| [11] | 李艳云, 王作敏. 大辽河口和辽东湾海域水质溶解氧与COD、无机氮、磷及初级生产力的关系[J]. 中国环境监测, 2006, 22(3): 70–72. Li Y Y, Wang Z M. The relation among dissolution oxygen (DO) to COD, inorganic nitrogen, reactive phosphate and primary Yield-power in the Liaodong gulf and seaport of daliaohe[J]. Environmental Monitoring in China, 2006, 22(3): 70–72. |
| [12] | 黄震, 余静. 长江芜湖段水体中溶解氧现状及其影响因素[J]. 安徽师范大学学报 (自然科学版), 2005, 28(3): 348–351. Huang Z, Yu J. The dissolved oxygen in water and influencing factors in the section of ChangJiang of Wuhu[J]. Journal of Anhui Normal University (Natural Science), 2005, 28(3): 348–351. |
| [13] | 罗献宝, 文军, 骆东奇, 等. 千岛湖水质变化特征与趋势分析[J]. 中国生态农业学报, 2006, 14(4): 208–212. Luo X B, Wen J, Luo D Q, et al. Changing characteristic and trend analysis of water quality of Qiandao Lake[J]. Chinese Journal of Eco-Agriculture, 2006, 14(4): 208–212. |
| [14] | 余员龙.千岛湖浮游植物群落结构时空分布格局及其与主要环境因子的关系[D].上海:上海海洋大学, 2010. Yu Y L. Spatial and temporal distribution of the phytoplankton community of lake Qiandaohu and its relationship with the main environmentaI factors[D]. Shanghai:Shanghai Ocean University, 2010. |
| [15] | 韩晓霞, 朱广伟, 吴志旭, 等. 新安江水库 (千岛湖) 水质时空变化特征及保护策略[J]. 湖泊科学, 2013, 25(6): 836–845. Han X X, Zhu G W, Wu Z X, et al. Spatial-temporal variations of water quality parameters in Xin'anjiang Reservoir (Lake Qi-andao) and the water protection strategy[J]. Journal of Lake Sciences, 2013, 25(6): 836–845. DOI: 10.18307/2013.0607 |
| [16] | 陈东, 张丽旭, 刘汉奇, 等. 长江口海域春夏季溶解氧分布特征及其相关因素分析[J]. 海洋环境科学, 2008, 27(S1): 49–53. Chen D, Zhang L X, Liu H Q, et al. Distribution characteristics and correlating factors analysis of dissolved oxygen in spring and summer in the Yangtze Estuary[J]. Marine Environmental Science, 2008, 27(S1): 49–53. |
| [17] | 刘流, 刘德富, 肖尚斌, 等. 水温分层对三峡水库香溪河库湾春季水华的影响[J]. 环境科学, 2012, 33(9): 3046–3050. Liu L, Liu D F, Xiao S B, et al. Effects of thermal stratification on spring blooms in Xiangxi Bay of the Three Gorges Reservoir[J]. Environmental Science, 2012, 33(9): 3046–3050. |
| [18] | 朱梦娟.梁子湖浮游植物群落结构及其与水环境理化因子的关系[D].武汉:华中农业大学, 2015. Zhu M J. Phytoplankton community structure and its relationship with environmental factors in Liangzhihu lake[D]. Wuhan:Huazhong Agricultural University, 2015. |
| [19] | 吴志旭, 刘明亮, 兰佳, 等. 新安江水库 (千岛湖) 湖泊区夏季热分层期间垂向理化及浮游植物特征[J]. 湖泊科学, 2012, 24(3): 460–465. Wu Z X, Liu M L, Lan J, et al. Vertical distribution of phytoplankton and physico-chemical characteristics in the lacustrine zone of Xin'anjiang Reservoir (Lake Qiandao) in subtropic China during summer stratifi-cation[J]. Journal of Lake Sciences, 2012, 24(3): 460–465. DOI: 10.18307/2012.0319 |
| [20] | 国家环境保护总局. 水和废水监测分析方法[M]. (第四版). 北京: 中国环境科学, 2002. |
| [21] | 胡鸿, 魏印心. 中国淡水藻类-系统、分类及生态[M]. 北京: 科学出版社, 2006. |
| [22] | Monterey G I, Dewitt L M. Seasonal Variability of Global Mixed Layer Depth from WORD 98 Temperature and Salinity Profiles[M]. Pacific Grove, California: National Oceanic and Atmospheric Administration, 2000. |
| [23] | 李媛, 刘德富, 孔松, 等. 三峡水库蓄泄水过程对香溪河库湾水华影响的对比分析[J]. 环境科学学报, 2012, 32(8): 1882–1893. Li Y, Liu D F, Kong S, et al. Comparative analysis of the impact of discharging and impounding process of Three Gorges Reservoir on the algal bloom in the Xiangxi Bay[J]. Acta Scientiae Circumstantiae, 2012, 32(8): 1882–1893. |
| [24] | Reinart A, Arst H, Erm A, et al. Optical and biological properties of Lake ülemiste, a water reservoir of the city of Tallinn Ⅱ:light climate in Lake ulemiste[J]. Lakes & Reservoirs:Research and Management, 2001, 6(1): 75–84. |
| [25] | 董春颖, 虞左明, 吴志旭, 等. 千岛湖湖泊区水体季节性分层特征研究[J]. 环境科学, 2013, 34(7): 2574–2581. Dong C Y, Yu Z M, Wu Z X, et al. Study on seasonal characteristics of thermal stratification in lacustrine zone of lake Qiandao[J]. Environmental Science, 2013, 34(7): 2574–2581. |
| [26] | 盛海燕, 虞左明, 韩轶才, 等. 亚热带大型河流型水库——富春江水库浮游植物群落及其与环境因子的关系[J]. 湖泊科学, 2010, 22(2): 235–243. Sheng H Y, Yu Z M, Han Y C, et al. Phytoplankton community and its relationship with environmental factors of a large riverine reservoir, Fuchunjiang Reservoir in subtropical China[J]. Journal of Lake Sciences, 2010, 22(2): 235–243. |
| [27] | 曾艳艺, 黄翔鹄. 温度、光照对小环藻生长和叶绿素a含量的影响[J]. 广东海洋大学学报, 2007, 27(6): 36–40. Zeng Y Y, Huang X H. Effects of temperature and illumination on growth and Chlorophyll-a of cyclotella sp[J]. Journal of Guangdong Ocean University, 2007, 27(6): 36–40. |
| [28] | 代玲玲.三峡水库富营养化藻类特征及环境因素影响研究[D].重庆:重庆大学, 2007. Dai L L. Study on euthophication algal characters and environmental factors effect of three Gorge Reservoir[D]. Chongqing:Chongqing University, 2007. |
| [29] | 石英, 童国榜, 王苏民, 等. 云南鹤庆盆地晚第四纪硅藻植物群与环境[J]. 海洋地质与第四纪地质, 1998, 18(4): 79–86. Shi Y, Tong G B, Wang S M, et al. Late quaternary diatom community and environment changes in Heqing basin, Yunnan Province[J]. Marine Geology & Quaternary Geology, 1998, 18(4): 79–86. |
| [30] | Raven J A, Geider R J. Temperature and algal growth[J]. New Phytologist, 1988, 110(4): 441–461. DOI: 10.1111/nph.1988.110.issue-4 |
| [31] | Eppley R W. Temperature and phytoplankton growth in the sea[J]. Fishery Bulletin, 1972, 70(4): 1063–1085. |
| [32] | 夏品华, 李秋华, 林陶, 等. 贵州高原百花湖水库湖沼学变量特征及环境效应[J]. 环境科学学报, 2011, 31(8): 1660–1669. Xia P H, Li Q H, Lin T, et al. Limnological characteristics and environmental effects of the Baihua Reservoir in GuiZhou plateau, China[J]. Acta Scientiae Circumstantiae, 2011, 31(8): 1660–1669. |
| [33] | 夏品华, 林陶, 李存雄, 等. 贵州高原红枫湖水库季节性分层的水环境质量响应[J]. 中国环境科学, 2011, 31(9): 1477–1485. Xia P H, Lin T, Li C X, et al. Features of the water column stratification and the response of water quality of Hongfeng Reservoir in Guizhou, China[J]. Chinese Journal of Environmental Science, 2011, 31(9): 1477–1485. |
| [34] | 邱华北, 商立海, 李秋华, 等. 水体热分层对万峰湖水环境的影响[J]. 生态学杂志, 2011, 30(5): 1039–1044. Qiu H B, Shang L H, Li Q H, et al. Impacts of seasonal thermal stratification on the water environment of Wanfeng lake[J]. Chinese Journal of Ecology, 2011, 30(5): 1039–1044. |
| [35] | 何剑波.千岛湖水温、溶解氧及叶绿素a垂向特征研究[D].杭州:浙江工业大学, 2014. He J B. Vertical characteristics of water temperature, dissolved oxygen and Chlorophyll a in Qiandaohu lake[D]. Hangzhou:Zhejiang University of Technology, 2014. |
| [36] | 张朝能. 水体中饱和溶解氧的求算方法探讨[J]. 环境科学研究, 1999, 12(2): 54–55. Zhang C N. Study on calculation method of saturation values of dissolved oxygen in waters[J]. Research of Environmental Sciences, 1999, 12(2): 54–55. |
| [37] | 杨丽娜, 李正炎, 张学庆. 大辽河近入海河段水体溶解氧分布特征及低氧成因的初步分析[J]. 环境科学, 2011, 32(1): 51–57. Yang L N, Li Z Y, Zhang X Q. Distribution characteristics of dissolved oxygen and mechanism of hypoxia in the upper estuarine zone of the Daliaohe River[J]. Environmental Science, 2011, 32(1): 51–57. |
| [38] | Foley B, Jones I D, Maberly S C, et al. Long-term changes in oxygen depletion in a small temperate lake:effects of climate change and eutrophication[J]. Freshwater Biology, 2012, 57(2): 278–289. DOI: 10.1111/j.1365-2427.2011.02662.x |
| [39] | Yin K D, Lin Z F, Ke Z Y. Temporal and spatial distribution of dissolved oxygen in the Pearl River Estuary and adjacent coastal waters[J]. Continental Shelf Research, 2004, 24(16): 1935–1948. DOI: 10.1016/j.csr.2004.06.017 |
| [40] | Boehrer B, Schultze M. Stratification of lakes[J]. Reviews of Geophysics, 2008, 46(2): RG2005. |
| [41] | 王雨春, 朱俊, 马梅, 等. 西南峡谷型水库的季节性分层与水质的突发性恶化[J]. 湖泊科学, 2005, 17(1): 54–60. Wang Y C, Zhu J, Ma M, et al. Thermal stratification and paroxysmal deterioration of water quality in a Canyon·Reser·voir, Southwestern China[J]. Journal of Lake Sciences, 2005, 17(1): 54–60. DOI: 10.18307/2005.0109 |
| [42] | 游亮, 崔莉凤, 刘载文, 等. 藻类生长过程中DO、pH与叶绿素相关性分析[J]. 环境科学与技术, 2007, 30(9): 42–44. You L, Cui L F, Liu Z W, et al. Correlation analysis of parameters in algal growth[J]. Environmental Science & Technology, 2007, 30(9): 42–44. |
| [43] | 杨正健.分层异重流背景下三峡水库典型支流水华生消机理及其调控[D].武汉:武汉大学, 2014. Yang Z J. The mechanisms of algal blooms and its operation method through water level fluctuation under the situation of the bidirectional density currents in tributaries of the Three Gorges Reservior[D]. Wuhan:Wuhan University, 2014. |
| [44] | Kalff J.湖沼学-内陆水生态系统[M].古滨河, 刘正文, 李宽意, 译.北京:高等教育出版社, 2011. Kalff J. Limnology-Inland water ecosystems[M]. Gu B H, Liu Z W, Li K Y, Trans. Beijing:Higher Education Press, 2011. |
| [45] | 杨丽丽.千岛湖浮游动物群落结构特征及其与环境因子的关系[D].上海:上海海洋大学, 2012. Yang L L. Zooplankton community structure and its relation to environment factors in Qiandao Lake[D]. Shanghai:Shanghai Ocean University, 2012. |
| [46] | 朱爱娇, 姚建良, 薛俊增. 千岛湖蚤状溞垂直分布格局及其季节与昼夜变化[J]. 海洋湖沼通报, 2007(4): 120–128. Zhu A J, Yao J L, Xue J Z. The variation of the seasonal and diel vertical distribution of daphnia pulex in lake Qiandao[J]. Transactions of Oceanology and Limnology, 2007(4): 120–128. |
| [47] | 王先云, 王丽卿, 盖建军, 等. 千岛湖浮游甲壳动物垂直分布与昼夜垂直移动[J]. 动物学杂志, 2010, 45(5): 95–104. Wang X Y, Wang L Q, Gai J J, et al. Vertical distribution and diel vertical migration of planktonic crustaceans in Qiandao Lake[J]. Chinese Journal of Zoology, 2010, 45(5): 95–104. |
| [48] | 李共国, 虞左明. 浙江千岛湖浮游动物群落多样性研究[J]. 生物多样性, 2001, 9(2): 115–121. Li G G, Yu Z M. A study on the diversity of a zooplankton community in Lake Qiandaohu, Zhejiang[J]. Biodiversity Science, 2001, 9(2): 115–121. |
| [49] | 王庆安, 黄时达, 孙铁珩. 多藻浅水体中pH值和溶解氧协同周期性变化初探[J]. 四川环境, 2001, 20(2): 4–7, 29. Wang Q A, Huang S D, Sun T H. Study on the coordinate periodic change and the relativity between pH and DO in Shallow Water with algae[J]. Environment of Sichuan, 2001, 20(2): 4–7, 29. |
| [50] | 黄岁樑, 臧常娟, 杜胜蓝, 等. pH、溶解氧、叶绿素a之间相关性研究Ⅰ:养殖水体[J]. 环境工程学报, 2011, 5(6): 1201–1208. Huang S L, Zang C J, Du S L, et al. Study on the relationships among pH, dissolved oxygen and chlorophyll a Ⅰ:aquaculture water[J]. Chinese Journal of Environmental Engineering, 2011, 5(6): 1201–1208. |
| [51] | 陈洋, 杨正健, 黄钰铃, 等. 混合层深度对藻类生长的影响研究[J]. 环境科学, 2013, 34(8): 3049–3056. Chen Y, Yang Z J, Huang Y L, et al. Research on the influence of mixing layer depth on algal growth[J]. Environmental Science, 2013, 34(8): 3049–3056. |
| [52] | 杨东方, 高振会, 王培刚, 等. 营养盐Si和水温影响浮游植物的机制[J]. 海洋环境科学, 2006, 25(1): 1–6. Yang D F, Gao Z H, Wang P G, et al. Mechanism of effect of nutrient silicon and water temperature on phytoplankton[J]. Marine Environmental Science, 2006, 25(1): 1–6. |
| [53] | Chapman D V. Water quality assessments:a guide to the use of biota, sediments and water in environmental monitoring[M]. (2nd ed). Melbourne Madras: Chapman & Hall, 1996. |
| [54] | 王瑞巧, 陆宝宏, 王春燕, 等.新安江水库冬季水温分布结构及规律初探[A].见:第五届中国水论坛论文集[C].南京:中国水利学会, 中国自然资源学会, 2007. 377-382. |
| [55] | 田泽斌, 刘德富, 姚绪姣, 等. 水温分层对香溪河库湾浮游植物功能群季节演替的影响[J]. 长江流域资源与环境, 2014, 23(5): 700–707. Tian Z B, Liu D F, Yao X J, et al. Effect of water temperature stratification on the seasonal succession of phytoplankton function grouping in Xiangxi Bay[J]. Resources and Environment in the Yangtze Basin, 2014, 23(5): 700–707. |