 2017, Vol. 38
2017, Vol. 38



高含固[进料含固率(total solids,TS)>10%]厌氧消化是解决我国脱水污泥处理处置问题的重要途径.相比传统的低含固厌氧消化(含固率为1%~5%)[1],高含固厌氧消化技术甚至可以直接利用污水处理厂的脱水污泥(含固率为20%),因而具有设施体积小、水耗及能耗较低等优势.另外,高含固厌氧消化系统和传统的低含固厌氧消化系统相比,VS降解率基本相当,而高含固厌氧消化系统的单位容积产气率更高[2],具有更好的应用前景.
然而,由于污泥高含固厌氧消化系统的进料中含氮物质如蛋白质、尿素等含量更高,经过微生物的作用,它们会最终降解为小分子的氨氮并累积在系统中.过高的氨氮浓度会对微生物产生毒害作用[3],从而降低整个消化系统的产气性能.总氨氮(TAN)又分为铵根离子和游离态的氨(FAN),其中游离氨具有较高的细胞膜渗透性而被认为是产生氨抑制的主要原因[4].微生物被氨氮抑制的可能机制是游离氨作为疏水性分子,通过被动扩散作用进入细胞,破坏了细胞内外质子平衡并导致钾的缺乏[5].另外,进入细胞的游离氨在细胞内转变为铵,铵在细胞内积累改变了细胞内的pH,从而对细胞产生毒害作用[6].关于游离氨对厌氧消化系统的影响,由于系统的含固率、基质、温度和接种泥驯化程度等各种因素的不同,所得出的游离氨抑制阈值也从几十到几百毫克升不等[7].例如,传统的低含固厌氧消化系统中,FAN在145 mg·L-1,pH为7.43时便会使家禽粪便悬浮液厌氧消化的产气量下降27%[8].在高含固厌氧消化系统中,Hidaka等[9]的实验发现FAN在不到600 mg·L-1的条件下,高含固系统COD降解率就受到明显抑制.然而,微生物对于游离氨的耐受度可以更高,Kim等[10]发现FAN为700 mg·L-1时系统仍能稳定运行不受抑制.而Calli等[11]利用人工配水作为基质,在提高游离氨的过程中,COD降解率一直维持在78%~96%,即使在FAN最终达到800 mg·L-1时依然维持在80%,系统性能并没有受到抑制.所以,厌氧消化氨抑制研究的重点不仅在于获得游离氨的抑制阈值浓度,还在于研究游离氨浓度改变背后相关的微生物种群结构和代谢途径的变化,从而真正揭示氨抑制的机制.游离氨的计算方法与总氨氮、pH和温度有关,在温度不变的情况下,环境中的pH值越低,FAN也越低.例如在厌氧消化温度为37℃的条件下,当pH从8.0降低至7.0时,FAN占TAN的比例将会从11%下降至1.2%,因而降低pH是一种降低系统中FAN浓度的方法.不同的微生物的适宜pH范围也有所不同, 产甲烷菌的最适宜pH范围为6.5~7.8[12],而产酸菌最佳pH在5.5~7.0[3],控制pH在微生物适宜的范围内可能可以减轻FAN的抑制作用[9].
本实验采用9 L的半连续厌氧消化装置,以脱水污泥为原料,分别通过降低pH和人为投加氨氮的手段来降低和提高反应器内的游离氨浓度,以考察两种调控手段下游离氨浓度的变化对厌氧消化系统性能的影响.与此同时,利用高通量测序和定量PCR的分析方法,研究高含固污泥厌氧消化系统在不同的游离氨调控策略下性能参数以及相应的细菌和古菌种群结构和数量的变化,以期为最终合理控制高含固厌氧消化体系中游离氨浓度,保证消化系统良好稳定地运行提供理论支撑.
1 材料与方法 1.1 厌氧消化实验实验中的脱水污泥取自上海某污水处理厂的脱水污泥机房,脱水后的污泥含固率(TS)和挥发性固体(VS)占总固体比例(VS/TS)分别为20.1%和53.9%.实验所用接种污泥取自实验室中稳定运行的脱水污泥中温厌氧消化反应器,TS和VS/TS分别为13.2%和46.0%.进料脱水污泥和接种泥的碳氮比(C/N)分别为6.9:1和5.2:1.
半连续式反应器的有效容积为9 L,通过螺带式搅拌装置进行搅拌,内置的水浴温控装置能维持反应器内物料温度为(37±1)℃.每日产气量通过湿式气体流量计测定.反应器的转速为60 r·min-1,以转动/停止为10 min/10 min方式交替运行.运行的首日向反应器内加入4 kg接种泥和1 kg基质原泥,之后连续4 d每天添加1 kg事先通过自来水配制好的含固率为15%的脱水污泥至反应体系的总体积达到9 L,从第6 d开始按照固体停留时间(solid retention time,SRT)为20 d以半连续方式进出料(每天进出料各一次).
本实验共运行2台反应器(R1和R2). R1在稳定运行至35 d后每天将适量的浓HCl与进料混合,在第50 d反应器pH下降至7.09±0.02,随后稳定该pH值41 d,第92 d停止添加HCl直至实验结束. R2在稳定运行至56 d后,开始在进料中添加30 g NH4Cl,并依据TAN分析结果分别于第58 d和第67 d加入13 g和35 g NH4Cl以使得反应器内的TAN从3 284 mg·L-1增加到5 500 mg·L-1.从第70 d开始,为了维持TAN在5 000~6 000 mg·L-1,根据出料中氨氮的流失量计算,每天在进料中加入5 g NH4Cl至实验结束. 70~124 d这一阶段FAN从稳定阶段的(400±173) mg·L-1提高到了(526±25) mg·L-1.系统FAN浓度范围最终稳定在450~600 mg·L-1(总氨氮浓度为5 000~6 000 mg·L-1)至实验结束.
1.2 指标分析方法平均每周采集两次反应器的出料进行分析,测定pH、TS、VS/TS,挥发性脂肪酸(volatile fatty acids,VFAs)浓度、总碱度、总氨氮(TAN)浓度、气体成分.每天记录产气量. pH测定采用实验室用pH计(S210, METTLER, Switzerland).消化污泥样品在13 000 r·min-1下离心20 min后进行过滤,滤液经微孔滤膜(0.45 μm)过滤后,稀释一定倍数并加入甲酸调整pH≤2.0,再使用气相色谱[2010 plus, Shimadzu, Japan,火焰离子化检测器(FID)]测定VFA浓度.所使用的色谱柱为Rtx-WAX型毛细管柱(30 m×0.25 mm×0.25 μm). TS、VS/TS、总碱度和TAN浓度采用标准方法测得[13].沼气中甲烷和二氧化碳的含量测定采用装有热导检测器(TCD)的气相色谱仪(GC112A, INESA, China),色谱柱为GDX-102(2 m×4 mm).游离氨浓度根据总氨氮浓度、pH和温度计算得到[14], 具体计算公式如下:
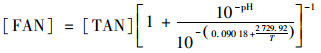
|
(1) |
式中,[FAN]和[TAN]分别为游离氨和总氨氮的浓度(mg·L-1);T为热力学温度(K).
VS降解率(VSreduction)由式(2)[15]计算得出(假定系统中无机组分,即不能被降解的物质的量保持恒定):
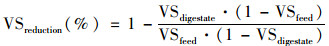
|
(2) |
式中, VSdigestate和VSfeed分别为厌氧消化后的沼渣和进料污泥中的VS占TS的比例(VS/TS,%).
1.3 微生物分析 1.3.1 微生物种群结构分析定期保留反应器出料于2 mL冻存管中,于-80℃冷冻保存. DNA样品使用土壤DNA提取试剂盒(PowerSoil,MO BIO, USA)提取.分析中选用的8个R1样品来自第28、35、42、49、77、88、109和124 d,代表了稳定、调控、抑制稳定和恢复这4个阶段.所选用的16个R2样品来自第4、10、16、21、28、35、42、49、56、58、63、70、77、84、91和102 d,代表了R2启动、稳定、提高游离氨、高游离氨抑制稳定这4个阶段.提取出的DNA经过PCR扩增,采用高通量测序(MiSeq4000, Illumina)分析其中古菌与细菌的种群结构. PCR产物先利用QuantiFLuorTM系统(Promega)测定浓度,随后依据AxyPrep DNA试剂盒(AXYGEN, USA)的凝胶回收的方法进行提纯.提纯的PCR产物的质量通过凝胶电泳确定.引物设计根据Illumina公司(San Diego, California, USA)的操作手册,针对细菌和古菌的16S rRNA基因的通用扩增引物对分别为338f (5′-ACTCCTACGGGAGGCAGCA-3′)/806r (5′-GGA CTACHVGGGTWTCTAAT-3′)[16]和344f (5′-ACGGG GYGCAGCAGGCGCGA-3′)/915r (5′-GTGCTCCCCC GCCAATTCCT-3′)[17].
1.3.2 甲烷菌定量PCR分析本研究中根据多样性分析结果,选取产甲烷菌中主要菌群甲烷八叠球菌科(Methanosarcinaceae),对其16S rRNA基因浓度变化进行了定量分析.定量PCR实验采用Sybergreen荧光染料法,引物设计采用甲烷八叠球菌科16S rRNA基因的特异引物对:前引物(5′-GAAACCGYGATAAGGGGA-3′)和后引物(5′-TAGCGARCATCGTTTACG-3′)[18].使用定量PCR仪(7500 Real Time PCR System,Applied Biosystems)对Methanosarcinaceae进行定量分析. Sybergreen定量PCR反应体系为10 μL SYBR Premix Ex TaqⅡ (Tli RNaseH Plus) (2x) (TaKaRa, Japan), 0.8 μL前引物(10 μmol·L-1), 0.8 μL后引物(10 μmol·L-1), 0.4 μL ROXⅡ,2 μL模板DNA和PCR等级无菌水,最终体积为20 μL.每个样品3个平行.反应步骤:先95℃下预变性30 s,然后经过95℃下5 s, 55℃下30 s和72℃下40 s循环40次.每一次循环的最后一个步骤检测荧光值.
2 结果与讨论 2.1 游离氨调控对系统性能的影响 2.1.1 降低游离氨浓度对产气量和VS降解率的影响R1的相关理化指标变化曲线如图 1所示.在反应器启动阶段(0~24 d),日产气量波动较大,并且在第13 d和第24 d出现两个产气高峰.该阶段有大量的VFA积累,TAN含量也很高,说明启动阶段水解和产酸菌活性相对于产甲烷菌活性更高,能够快速降解大量的大分子有机物并生成VFA,而此时产甲烷菌由于其世代周期相对较长,还不能大量增殖并及时消耗乙酸,导致VFA积累和产气剧烈波动. 25 d后,R1的产气量逐渐稳定,pH也稳定在7.98±0.04,稳定阶段(25~35 d) FAN约为(329±40) mg·L-1. Braun等[19]利用液态的猪粪进行厌氧发酵,提出FAN在150 mg·L-1时系统就开始受到抑制,他们的反应系统在降低pH之后获得了更好的产气量.同样,Zeeman等[20]在牛粪的高温厌氧消化过程中,TAN为3 000 mg·L-1,发现在pH为7.5时,性能很差,而当他们将pH降到7.0后,由于FAN的降低,甲烷的产量增长了4倍.因此,为了使得游离氨浓度下降,减轻微生物所受的游离氨的抑制作用,获得更好的产气性能,从第35 d开始人为地向R1中添加HCl,使得pH逐渐下降并在50 d之后稳定在7.09±0.02.尽管FAN在50~91 d从之前的(329±40) mg·L-1下降到了(47±13) mg·L-1,但是这个阶段的日产气量平均为(10.8±0.3) L·d-1,相比稳定阶段的(14.4±1.1) L·d-1却减少了25%.由表 1可知,VS降解率也和日产气量相似,随着游离氨浓度下降而降低,并未出现与之前的研究[19, 20]相一致的结果.在第91 d停止添加HCl之后,pH和FAN逐渐恢复到厌氧消化的初始范围,VS降解率和日产气量也随之上升.可见,在本实验的高含固厌氧消化系统中,降低pH虽然明显降低了FAN的浓度,但是并没有达到预期的减轻游离氨对微生物的毒害作用进而提升系统产气性能的目的.相反,从pH下降和回升的两个阶段的数据都可看出,pH的降低明显影响了系统有机物的水解率和VS降解率,进而导致产酸菌和产甲烷菌的底物供给不足,最终引起产气量的下降.

|
图 1 R1的性能参数随时间的变化曲线 Fig. 1 Time course of operational parameters of R1 during the operation |
|
|
表 1 R1在不同阶段的各项指标(置信区间95%) Table 1 Summarized operational parameters of R1 during the four phases (with 95% confidence intervals) |
2.1.2 提高游离氨浓度对产气量和VS降解率的影响
与R1反应器不同,R2是通过提高TAN的浓度来调控FAN的浓度.由图 2可知,R2在启动阶段(0~39 d)和R1相似,产气量波动较大,在第15 d和第26 d分别出现产气高峰. R2在40 d之后达到稳定,稳定阶段(40~56 d)日产气量为(12.0±0.3) L·d-1,VS降解率为(34.1±1.1)%,甲烷含量约为64.8%,各项性能参数与之前稳定运行的污泥高含固厌氧消化反应器接近[21].由图 2和表 2可知,随着FAN的提升,日产气量和VS降解率都出现了明显下降,相比稳定阶段分别减少了14.2%和33.7%.虽然外加氨氮之后,由于NH4Cl与污泥中的OH-能进行反应,导致在提升氨氮初期H+浓度增加,pH有所下降.但是,pH的变化幅度较小,70 d以后系统pH又逐渐恢复,尽管游离氨浓度超过500 mg·L-1,但消化系统仍然能保持稳定运行,说明系统处于“抑制的稳定状态”[3].

|
图 2 R2的性能参数随时间的变化曲线 Fig. 2 Time course of operational parameters of R2 during the operation |
|
|
表 2 R2在不同阶段的各项指标(置信区间95%) Table 2 Summarized operational parameters of R2 during the four phases (with 95% confidence intervals) |
2.2 游离氨调控对细菌种群的影响
当通过降低pH的方式来降低R1系统中游离氨浓度时,细菌种群的变化如图 3所示.从中可知,Candidate_division_WS 6 _norank在恢复阶段含量明显提高.然而,已有研究中尚未记载这种细菌的具体功能,因而也无法判断其在系统中所起的作用.

|
图 3 R1中细菌相对丰度随时间的变化 Fig. 3 Variations of the bacterial relative abundance of R1 during the operation |
另一方面,在前两个阶段中,相对丰度占前几位的分别为OPB 54 _norank(14%~26%)、Proteiniphilum(7%~11.7%)、Family_XIV_uncultured(5%~13%)和D8A-2_norank(1%~3.9%,其中第49 d占13.2%),在抑制稳定阶段这些微生物的相对丰度全部都下降到5%以下,显然随着pH条件的改变,这些细菌的生长和代谢受到了影响. Proteiniphilum是一种产酸菌,主要产物是乙酸和丙酸,能够进行解朊作用降解蛋白质并产生NH3[22].本实验中,相对丰度随pH下降幅度最大的细菌是OPB54 _norank. Hao等[23]利用乙酸为基质进行高温厌氧消化,并设置了低TAN (2 600 mg·L-1)和高TAN (7 000 mg·L-1)两组实验,发现OPB54(具体功能未知)在两种氨氮浓度下均是含量最多的细菌,但是在低FAN下相对丰度更高,这与本研究的结果并不一致.除此之外,一些第一阶段含量较少的细菌如Tepidimicrobium(3.9%)、Macellibacteroides(1.8%)、Caldicoprobacter(1.5%)也都随着pH调控游离氨下降而种群丰度明显降低. Tepidimicrobium能分解葡萄糖等多种糖类,也可以降解蛋白类的物质,产物为乙酸、乙醇、H2和CO2等,有研究表明Tepidimicrobium最适宜的pH范围是8~8.5[24, 25],本实验中pH降低到7.1可能会是抑制该菌属生长的一个原因.同样情况下,由于Macellibacteroides和Caldicoprobacter是能分解糖类的细菌,他们的最终代谢产物也为小分子的VFA、H2和CO2等[26, 27],这两类细菌丰度的下降也在一定程度上解释了有机物含量升高的原因.由此可见,通过pH调控游离氨浓度使得系统理化环境不适宜于原本一些丰度较大的降解糖类和蛋白的产酸菌的生长,从而使得VS降解率下降.
由图 3可知,随着pH的调控,在抑制稳定阶段,理研菌属(Rikenellaceae_RC9 _gut_group)和Family_XIV_uncultured的相对丰度从之前的不足1%分别上升至24.3%和22.9%,Petrimonas和Acholeplasma也从不足1%分别增长到1.1%和4.9%.这其中,理研菌属Rikenellaceae不仅可以分解碳水化合物而且还可以降解蛋白质[28].而值得注意的是,随着游离氨浓度的上升恢复,这些微生物种群相对数量又明显下降,比如Rikenellaceae_RC9 _gut_group相对丰度又迅速下降到1%以下,Family_XIV_uncultured也降低至2.8%.有研究表明上述这些菌属虽然也都是进行糖类降解的产酸菌[29, 30],但从结果来看其代谢能力与Tepidimicrobium、Macellibacteroides和Caldicoprobacter相比明显较弱.互营单胞菌属Syntrophomonas的种群丰度在抑制稳定阶段也有明显增长,但在恢复阶段并没有下降,表明了这类微生物良好的pH适应范围.互营单胞菌属Syntrophomonas是一种互养型的具有氧化一些重要的中间产物如丙酸和丁酸的功能的细菌,代谢产物为乙酸和H2,该细菌必须要和能够消耗H2的微生物共同培养才能够生长[31],属于互养型的降解途径. Syntrophomonas在FAN升高时会受到抑制,在研究不同的工程规模[32]或者实验室规模[33]的厌氧反应器中的微生物种群结构时发现高游离氨浓度的反应器里例如Syntrophomonas、Pelotomaculum、Desulfobulbus等互养型微生物较低游离氨的反应器含量更少,显示互养型代谢更易受到游离氨的抑制.当为了降低游离氨浓度而控制pH值至7.0之后,系统中承担糖类和蛋白的降解的微生物类型发生了转化,之前丰度较大的该类细菌种群相对数量都出现降低,而另一些之前丰度较小的细菌含量增加,与此同时反应器的有机物降解性能和产气性能出现下降.
而当通过外加氨氮提高R2系统中游离氨浓度时,细菌种群的变化如图 4所示, R2反应器中3种主要的优势细菌种群,在启动阶段,Tepidimicrobium和Proteiniborus的相对丰度分别在第21 d和第16 d达到各自的最高值33.8%和15.8%.而后,两种细菌都在35 d下降,分别为18.3%和6.2%.细菌种群的丰度在稳定阶段保持基本不变,除了Anaerobranca在第42 d有一个激增.当第56 d开始,随着R2内游离氨浓度的不断上升,Anaerobranca也出现了明显的随之上升的趋势,而Tepidimicrobium和Proteiniborus则表现出随着FAN上升而降低的趋势. Tepidimicrobium是一种解胨型的细菌,可以以多种蛋白质的物质为基质来维持自己的生长[25],在R1和R2两组实验中,都发现了Tepidimicrobium的相对丰度随着游离氨的变化明显地改变,即不论是游离氨浓度提高还是降低,Tepidimicrobium的丰度都有所下降.和Tepidimicrobium类似,Proteiniborus也可以在pH值6~8的范围里利用蛋白质作为发酵底物[34]. Anaerobranca本质上也是分解蛋白质的细菌[35]. Prowe等[36]发现Anaerobranca只包含几种可以厌氧发酵蛋白转化为乙酸的厌氧微生物.本实验中第42 d Anaerobranca的增多同时也伴随着游离氨浓度的上升,可以看出当游离氨浓度变高的时候,Anaerobranca在整个蛋白的分解过程中所起的作用更大.而Anaerobranca对蛋白质的降解速率可能小于Tepidimicrobium和Proteiniborus,从而导致系统VS降解率的下降.

|
图 4 R2中3种优势细菌相对丰度随时间的变化 Fig. 4 Variations of the relative abundance of three dominant bacteria in R2 during the operation |
由图 5可知,R1反应器中游离氨浓度下降之后,氢利用型的甲烷囊菌属(Methanoculleus)的相对丰度明显降低,从13%下降到1.9%,并且在恢复阶段也未能完全恢复至原有水平,继续下降至不足1%.而整个过程中,乙酸型的甲烷八叠球菌属(Methanosarcina)相对丰度一直大于80%,在与甲烷囊菌属的竞争中,随着游离氨浓度和pH的下降,逐渐占据了更大的优势,最后丰度甚至达到97%.但是因为系统的产气量和VS降解率随着FAN和pH的下降而降低,为了更准确地观察游离氨及pH对产甲烷菌种群尤其是甲烷八叠球菌(Methanosarcina)的影响,通过定量PCR的分析而获得其16S rRNA基因浓度的变化趋势来定量分析甲烷菌种群的动态变化.由图 6知,R1反应器内相对丰度最大的甲烷八叠球菌的16S rRNA基因浓度(以湿泥计,下同)在第28 d和35 d分别为(1.27×106±8.0×104) copies·g-1和(1.19×106±9.89×104) copies·g-1,之后尽管pH不断降低,但依然维持在同一个数量级内,波动变化不大.而到了第88 d 16S rRNA基因浓度明显下降,为(1.27×105±1.22×104) copies·g-1,这时系统pH值已经在7.1维持了38 d,约2个SRT,显然之前pH为7左右的长时间作用已经使得较多的甲烷八叠球菌受到底物持续供给不足的影响.之后随着FAN和pH的回升,甲烷八叠球菌16S rRNA基因浓度又回升至106 copies·g-1的数量级水平.可见产甲烷菌种群在通过pH调控游离氨的过程中由于细菌种群结构的影响而受到基质供给不足的抑制,但随着pH的回升又较快恢复了活性.

|
图 5 R1的古菌在属的水平上随时间的变化 Fig. 5 Variations of relative abundance of archaea in R1 at genus level during the operation |

|
图 6 R1甲烷八叠球菌数量随时间的变化(n=3) Fig. 6 Changes in Methanosarcina 16S rRNA gene concentrations of R1(n=3) |
R2样品的古菌多样性分析结果如图 7所示,乙酸型的甲烷八叠球菌Methanosarcina为优势种群,在整个实验过程中占古菌的比例一直在92%以上,说明甲烷八叠球菌可能并没有受到游离氨浓度提高的抑制.定量PCR的结果也表明在游离氨浓度从(400±173) mg·L-1提高到(526±25) mg·L-1之后,R2内的甲烷八叠球菌的16S rRNA基因平均浓度从(3.43×106±2.91×106) copies·g-1变化为(3.58×106±1.18×106) copies·g-1,数量基本相同.因而,有机物降解受到抑制的主要原因应该来源于细菌种群结构所受的影响[37].

|
图 7 R2的古菌在属的水平上随时间的变化 Fig. 7 Variations of relative abundance of archaea in R2 at genus level during the operation |
(1)当FAN从(400±173) mg·L-1提高到(526±25) mg·L-1后,相比正常的消化状态,系统的日产气量和VS降解率分别降低了14.2%和33.7%,但依然能在抑制的状态下稳定运行.
(2)当通过pH调控使得FAN从(330±99) mg·L-1下降至(47±13) mg·L-1,系统的日产气量和VS降解率分别降低了25%和26.9%,而随着pH和FAN恢复至正常水平,系统的产气性能和VS降解率又恢复至原有水平,相比抑制稳定阶段分别提高了9.3%和31.6%.
(3)本文中的两种游离氨调控方式都会影响污泥高含固厌氧消化系统中细菌的种群结构.投加外来氨氮后,过高的FAN (500~600 mg·L-1)会使得降解蛋白的细菌种类朝着蛋白降解率更低的细菌菌群转变,但并没有影响到主要的产甲烷菌(甲烷八叠球菌)的生长;另一方面,通过降低pH来降低游离氨的浓度并不一定能使得性能提高,反而可能会由于pH的变化影响一些的糖类和蛋白质类发酵细菌的活性,导致水解效率的下降,引发后续产甲烷菌的底物供给不足,削弱系统产气性能.
| [1] | 胡玉瑛, 吴静, 王士峰, 等. 热处理对猪粪高固厌氧消化产甲烷能力的影响[J]. 环境科学, 2015, 36(8) : 3094–3098. Hu Y Y, Wu J, Wang S F, et al. Impact of thermal treatment on biogas production by anaerobic digestion of high-solid-content swine manure[J]. Environmental Science, 2015, 36(8) : 3094–3098. |
| [2] | 王广启, 吴静, 左剑恶, 等. 城市污泥高固体浓度厌氧消化的研究进展[J]. 中国沼气, 2013, 31(6) : 9–12. Wang G Q, Wu J, Zuo J E, et al. Progress of high-solid-content anaerobic digestion of municipal sludge[J]. China Biogas, 2013, 31(6) : 9–12. |
| [3] | Rajagopal R, Massé D I, Singh G. A critical review on inhibition of anaerobic digestion process by excess ammonia[J]. Bioresource Technology, 2013, 143 : 632–641. DOI: 10.1016/j.biortech.2013.06.030 |
| [4] | De Baere L A, Devocht M, Van Assche P, et al. Influence of high NaCl and NH4Cl salt levels on methanogenic associations[J]. Water Research, 1984, 18(5) : 543–548. DOI: 10.1016/0043-1354(84)90201-X |
| [5] | Sprott G D, Shaw K M, Jarrell K F. Ammonia/potassium exchange in methanogenic bacteria[J]. The Journal of Biological Chemistry, 1984, 259(20) : 12602–12608. |
| [6] | Sprott G D, Patel G B. Ammonia toxicity in pure cultures of methanogenic bacteria[J]. Systematic and Applied Microbiology, 1986, 7(2-3) : 358–363. DOI: 10.1016/S0723-2020(86)80034-0 |
| [7] | Chen Y, Cheng J J, Creamer K S. Inhibition of anaerobic digestion process:a review[J]. Bioresource Technology, 2008, 99(10) : 4044–4064. DOI: 10.1016/j.biortech.2007.01.057 |
| [8] | Webb A R, Hawkes F R. The anaerobic digestion of poultry manure:variation of gas yield with influent concentration and ammonium-nitrogen levels[J]. Agricultural Wastes, 1985, 14(2) : 135–156. DOI: 10.1016/S0141-4607(85)80025-1 |
| [9] | Hidaka T, Wang F, Togari T, et al. Comparative performance of mesophilic and thermophilic anaerobic digestion for high-solid sewage sludge[J]. Bioresource Technology, 2013, 149 : 177–183. DOI: 10.1016/j.biortech.2013.09.033 |
| [10] | Kim D H, Oh S E. Continuous high-solids anaerobic co-digestion of organic solid wastes under mesophilic conditions[J]. Waste Management, 2011, 31(9-10) : 1943–1948. DOI: 10.1016/j.wasman.2011.05.007 |
| [11] | Calli B, Mertoglu B, Inanc B, et al. Effects of high free ammonia concentrations on the performances of anaerobic bioreactors[J]. Process Biochemistry, 2005, 40(3-4) : 1285–1292. DOI: 10.1016/j.procbio.2004.05.008 |
| [12] | Rosenberg E, Delong E F, Lory S, et al. The prokaryotes[M]. Berlin Heidelberg: Springer, 2013. |
| [13] | APHA, AWWA, WEF. Standard methods for the examination of water and wastewater[M]. 21th ed.. Washington, DC: American Public Health Association, 2005. |
| [14] | Hansen K H, Angelidaki I, Ahring B K. Anaerobic digestion of swine manure:inhibition by ammonia[J]. Water Research, 1998, 32(1) : 5–12. DOI: 10.1016/S0043-1354(97)00201-7 |
| [15] | Koch K. Calculating the degree of degradation of the volatile solids in continuously operated bioreactors[J]. Biomass and Bioenergy, 2015, 74 : 79–83. DOI: 10.1016/j.biombioe.2015.01.009 |
| [16] | Lee C K, Barbier B A, Bottos E M, et al. The Inter-Valley soil comparative survey:the ecology of Dry Valley edaphic microbial communities[J]. The ISME Journal, 2012, 6(5) : 1046–1057. DOI: 10.1038/ismej.2011.170 |
| [17] | Gregory K B, Lovley D R. Remediation and recovery of uranium from contaminated subsurface environments with electrodes[J]. Environmental Science & Technology, 2005, 39(22) : 8943–8947. |
| [18] | Yu Y, Lee C, Kim J, et al. Group-specific primer and probe sets to detect methanogenic communities using quantitative real-time polymerase chain reaction[J]. Biotechnology & Bioengineering, 2005, 89(6) : 670–679. |
| [19] | Braun R, Huber P, Meyrath J. Ammonia toxicity in liquid piggery manure digestion[J]. Biotechnology Letters, 1981, 3(4) : 159–164. |
| [20] | Zeeman G, Wiegant W M, Koster-Treffers M E, et al. The influence of the total-ammonia concentration on the thermophilic digestion of cow manure[J]. Agricultural Wastes, 1985, 14(1) : 19–35. DOI: 10.1016/S0141-4607(85)80014-7 |
| [21] | Duan N N, Dong B, Bing W, et al. High-solid anaerobic digestion of sewage sludge under mesophilic conditions:feasibility study[J]. Bioresource Technology, 2012, 104 : 150–156. DOI: 10.1016/j.biortech.2011.10.090 |
| [22] | Chen S Y, Dong X Z. Proteiniphilum acetatigenes gen. nov., sp. nov., from a UASB reactor treating brewery wastewater[J]. International Journal of Systematic and Evolutionary Microbiology, 2005, 55(6) : 2257–2261. DOI: 10.1099/ijs.0.63807-0 |
| [23] | Hao L P, Lü F, Mazéas L, et al. Stable isotope probing of acetate fed anaerobic batch incubations shows a partial resistance of acetoclastic methanogenesis catalyzed by Methanosarcina to sudden increase of ammonia level[J]. Water Research, 2015, 69 : 90–99. DOI: 10.1016/j.watres.2014.11.010 |
| [24] | Niu L L, Song L, Liu X L, et al. Tepidimicrobium xylanilyticum sp. nov., an anaerobic xylanolytic bacterium, and emended description of the genus Tepidimicrobium[J]. International Journal of Systematic and Evolutionary Microbiology, 2009, 59(11) : 2698–2701. DOI: 10.1099/ijs.0.005124-0 |
| [25] | Slobodkin A I, Tourova T P, Kostrikina N A, et al. Tepidimicrobium ferriphilum gen. nov., sp. nov., a novel moderately thermophilic, Fe (Ⅲ)-reducing bacterium of the order Clostridiales[J]. International Journal of Systematic and Evolutionary Microbiology, 2006, 56(2) : 369–372. DOI: 10.1099/ijs.0.63694-0 |
| [26] | Jabari L, Gannoun H, Cayol J L, et al. Macellibacteroides fermentans gen. nov., sp. nov., a member of the family Porphyromonadaceae isolated from an upflow anaerobic filter treating abattoir wastewaters[J]. International Journal of Systematic and Evolutionary Microbiology, 2012, 62(10) : 2522–2527. |
| [27] | Yokoyama H, Wagner I D, Wiegel J. Caldicoprobacter oshimai gen. nov., sp. nov., an anaerobic, xylanolytic, extremely thermophilic bacterium isolated from sheep faeces, and proposal of Caldicoprobacteraceae fam. nov.[J]. International Journal of Systematic and Evolutionary Microbiology, 2010, 60(1) : 67–71. DOI: 10.1099/ijs.0.011379-0 |
| [28] | Su X L, Tian Q, Zhang J, et al. Acetobacteroides hydrogenigenes gen. nov., sp. nov., an anaerobic hydrogen-producing bacterium in the family Rikenellaceae isolated from a reed swamp[J]. International Journal of Systematic and Evolutionary Microbiology, 2014, 64(9) : 2986–2991. |
| [29] | Grabowski A, Tindall B J, Bardin V, et al. Petrimonas sulfuriphila gen. nov., sp. nov., a mesophilic fermentative bacterium isolated from a biodegraded oil reservoir[J]. International Journal of Systematic and Evolutionary Microbiology, 2005, 55(3) : 1113–1121. DOI: 10.1099/ijs.0.63426-0 |
| [30] | Tully J G, Whitcomb R F, Rose D L, et al. Acholeplasma brassicae sp. nov. and Acholeplasma palmae sp. nov., two non-sterol-requiring mollicutes from plant surfaces[J]. International Journal of Systematic Bacteriology, 1994, 44(4) : 680–684. DOI: 10.1099/00207713-44-4-680 |
| [31] | Mcinerney M J, Bryant M P, Hespell R B, et al. Syntrophomonas wolfei gen. nov. sp. nov., an anaerobic, syntrophic, fatty acid-oxidizing bacterium[J]. Applied and Environmental Microbiology, 1981, 41(4) : 1029–1039. |
| [32] | Li J B, Rui J P, Yao M J, et al. Substrate type and free ammonia determine bacterial community structure in full-scale mesophilic anaerobic digesters treating cattle or swine manure[J]. Frontiers in Microbiology, 2015, 6 : 1337. |
| [33] | Solli L, Håvelsrud O E, Horn S J, et al. A metagenomic study of the microbial communities in four parallel biogas reactors[J]. Biotechnology for Biofuels, 2014, 7 : 146. DOI: 10.1186/s13068-014-0146-2 |
| [34] | Niu L L, Song L, Dong X Z. Proteiniborus ethanoligenes gen. nov., sp. nov., an anaerobic protein-utilizing bacterium[J]. International Journal of Systematic and Evolutionary Microbiology, 2008, 58(1) : 12–16. DOI: 10.1099/ijs.0.65108-0 |
| [35] | Engle M, Li Y H, Woese C, et al. Isolation and characterization of a novel alkalitolerant thermophile, Anaerobranca horikoshii gen. nov., sp. Nov.[J]. International Journal of Systematic Bacteriology, 1995, 45(3) : 454–461. DOI: 10.1099/00207713-45-3-454 |
| [36] | Prowe S G, Antranikian G. Anaerobranca gottschalkii sp. nov., a novel thermoalkaliphilic bacterium that grows anaerobically at high pH and temperature[J]. International Journal of Systematic and Evolutionary Microbiology, 2001, 51(2) : 457–465. DOI: 10.1099/00207713-51-2-457 |
| [37] | Dai X, Yan H, Li N, et al. Metabolic adaptation of microbial communities to ammonium stress in a high solid anaerobic digester with dewatered sludge[J]. Scientific Reports, 2016, 6 : 28193. DOI: 10.1038/srep28193 |