 2017, Vol. 38
2017, Vol. 38



水陆交错带水生植物是湖泊水生态系统重要的初级生产者,是生态系统内的能量流动和物质循环的参与者,其群落结构与分布面积对湖泊水质有较大的影响[1].谢杰等[2]通过研究滇池、洱海水生植物与水质之间的关系,指出湖滨水生植物可以有效减少水体中氮磷营养盐,抑制藻类,减轻水体的富营养化.方焰星等[3]在室内水培试验中研究了石菖蒲、香菇草等水生植物对水体氮磷的去除效果,得出了水生植物具有良好去磷的效果和一定的固氮能力.有研究者指出,湖滨带底质是氮磷物质的聚集区域,构成了湖滨生态系统潜在的内源污染[4].在风浪条件下湖滨带底质上扬还会进一步恶化水体,释放氮磷[5].湖滨带水生植物群落通过对底质中氮磷物质的同化、迁移,对水体富营养化控制,浮游生物群落稳定起到一定作用[6].王磊等[7]通过研究太湖湖滨带表层沉积物氮磷与挺水植物芦苇、茭白分布情况的相关关系,指出了两种水生植物可以通过根部有效吸收、固定氮磷. Horppila等[8]在芬兰南部希登韦西湖(Lake Hiidenvesi)研究指出湖滨内源磷荷载与悬浮颗粒物之间的关系受到沉水植物的影响,在沉水植物区内,表层底质含磷量较低,其上扬的悬浮颗粒物可以从水体中吸收溶解性磷,减少区域内上覆水的溶解性磷浓度. Özbay[9]在地中海土耳其境内的卡里湖(Cali Lake)进行了野外水质监测,指出了卡里湖湖滨带的水质状况及底质的悬浮情况受到两栖蓼和狐尾藻的直接影响.目前研究者已经指出湖滨带水生植物在抑制底质氮磷释放方面具有的重要作用,尤其是在风浪影响下湖滨带植物对底质氮磷释放的抑制作用更加明显.本文通过选择5种常见水陆交错带植物对底质改进,研究其在模拟风浪条件下抑制底质氮磷释放的过程和效果;同时分析影响这种抑制作用的主要因素,比较几种植物的抑制效果,以期为相关的研究和工程应用提供参考.
1 材料与方法 1.1 植物与底质来源试验所用的植物和部分底质土样来自于无锡贡湖湾太湖新城贡湖生态修复示范工程区(N 31°27′19.91″~31°27′25.38″、E 120°19′48.04″~120°19′56.39″),该示范区原为鱼塘,2013年退渔还湖后经过基底修复改善,于2014年5月经人工植被修复后形成水陆交错带示范区.示范区内底质主要为外来覆土,属于湖滨带生态修复区域新生底质,其孔隙度37%,含水率24%,粒径分布以100 μm以下的细小颗粒为主.试验中还采集生态修复示范区外的沙壤底质作为试验对象,其孔隙度45%,含水率25%,粒径分布以100 μm以上的中细、中粗颗粒为主.
试验中水陆交错带植物均为长江中下游常见种类,于2014年5月人工种植后生长稳定并形成群落.采集示范区分布面积大、生长良好的5种水生植物,包括李氏禾(Leersia hexandra Swartz)、双穗雀稗(Paspalum distichumL.)、香菇草(Hydrocotyle vulgaris)、黄菖蒲(Iris pseudacorus L.)、黄花水龙(Ludwigia peploides)作为试验对象[10~14].
1.2 试验设置本试验装置经塑料养殖箱改造而成,如图 1所示,每个养殖箱底均匀铺设20 cm底质. 5种水陆交错带植物分别在示范工程区新生底质和沙壤底质两种底质中进行种植,设置一组平行样,两种底质分别设置空白对照组.各组装置内水生植物栽种情况如表 1所示,各组植物的栽种生物量在0.5~1.1 kg·m-2,植物的株高株数因不同植物种类而存在差异.试验开始前5个试验组、1个空白组在植物栽种后加少量水,保持底质润湿,等待24 h观察栽种效果,确保试验植物与底质紧密结合.之后各组注入纯净水,淹没植物根部至水深10 cm.经过一周的生长适应期后开始试验,每个装置中放置曝气头,曝气量25 mL·s-1, 装置中上覆水流速1.9~2.5 m·s-1, 水力梯度15 s-1.

|
图 1 植物栽种试验装置示意 Fig. 1 Diagram of experimental devices |
|
|
表 1 各试验组设置基本参数 Table 1 Parameters of five experimental groups and blank control group |
1.3 数据采集与分析
试验开始后用曝气头模拟风浪,连续扰动至7 d监测上覆水悬浮物颗粒物浓度SS、高锰酸盐指数、总氮及总磷等指标;试验进行至14 d停止水力扰动,静止6 h后测试上述上覆水水质指标;继续水力扰动,在扰动15 d、20 d对上覆水SS、高锰酸盐指数、TN及TP等指标进行测定. 20 d试验结束时取出植物,用水清洗植物根部,筛选 < 1 mm须根,测定根长密度、根表面积密度与根重密度.
利用钼酸盐比色法测定水体总磷(total phosphorus),碱性过硫酸钾比色法测定水体总氮(total nitrogen),利用0.45 μm滤膜过滤的方式测定水体悬浮颗粒物浓度(suspended solid),利用高锰酸盐氧化法测定水体高锰酸盐指数.本试验中底质采用凯式定氮仪测定底质总氮,利用高氯酸-硫酸消解法测定底质总磷与有效磷.利用激光粒度仪测定底质粒径分布情况.试验数据利用EXCEL、Origin等软件进行整理,利用SPSS数学统计软件进行数据间相关性分析.底质氮、磷释放速率为上覆水中总氮、总磷增量百分比与时间的比值,公式为:
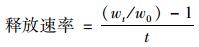
|
式中,w0为初始水体中的氮磷浓度,wt为试验运行t时间后水体中氮磷浓度,t为试验运行时间.试验中生物量采用试验前后测定其植物鲜重来表示,利用LA-S根系分析仪测定根长密度、根表面积密度与根重密度.
2 结果与讨论 2.1 植物生长对底质中氮释放影响 2.1.1 上覆水TN、NH4+-N、NO3--N变化如图 2中所示空白组上覆水TN、NH4+-N、NO3--N浓度在试验开始前两周持续升高,TN浓度由0.2 mg·L-1上升至0.45 mg·L-1, NH4+-N浓度由0.1 mg·L-1上升至0.2 mg·L-1,NO3--N在第14 d达到0.1 mg·L-1,15 d后TN、NH4+-N、NO3--N浓度略有下降.与空白组相比,各试验组TN浓度在试验开始时就较高,李氏禾组达到0.5 mg·L-1,黄菖蒲组达到0.4 mg·L-1,其他各组均超过0.2 mg·L-1,随后的两周试验中各组TN、NH4+-N、NO3--N浓度也持续升高,各组TN、NH4+-N、NO3--N浓度在第14 d或第15 d达到最高,随后有所下降.在各试验组之间,李氏禾组TN、NH4+-N、NO3--N浓度最高,其次为黄菖蒲组,TN、NH4+-N、NO3--N浓度稍低的为双穗雀稗、香菇草、黄花水龙试验组. DIN是上覆水中NO3--N和溶解性NH4+-N的总和,DIN/TN指示了可溶性无机氮占水体中TN的比例.如图 2(a)所示,空白组的DIN/TN比例在试验周期前两周扰动过程中由0.5上升至0.6,第14 d停止扰动期间DIN/TN比例下降,恢复扰动后比例也恢复至0.6.在各试验组中,双穗雀稗、香菇草、黄花水龙与空白组相反,在扰动过程中DIN/TN比例下降,停止扰动DIN/TN比例上升;而李氏禾、黄菖蒲DIN/TN比例在试验周期内持续升高.各组的DIN/TN比例变化差异说明曝气扰动带来的溶解氧变化不是影响上覆水中TN、NH4+-N、NO3--N浓度的主要因素,但水体扰动确实带来了上覆水中TN、NH4+-N、NO3--N浓度升高.各试验组TN、NH4+-N、NO3--N浓度变化差异说明了李氏禾、香菇草、黄花水龙的生长会带来底质氮释放的增加.

|
14(S)表示第14d停止跳动,下同 图 2 各试验组释放过程中上覆水TN、NH4+-N、NO3--N的浓度及比例变化情况 Fig. 2 Concentration and percentage of different forms of nitrogen (TN, NH4+-N, NO3--N) in the process of release |
通过计算试验期间各组底质向上覆水释放总氮的速率,可以发现差异主要在不同植物之间,SPSS软件双侧T检验结果显示同种植物组内两种底质间的总氮释放没有显著差异(α < 0.05).如图 3所示,空白对照组底质对上覆水TN的释放速率一直保持在0.02 d-1, 黄菖蒲试验组的释放速率与空白组十分接近,试验期间等于或低于0.02 d-1;双穗雀稗、黄花水龙试验组在扰动至14 d时达到最大释放率0.08 d-1,随后持续下降,双穗雀稗组释放率下降至0.02 d-1,黄花水龙试验组下降至0.045 d-1;香菇草、李氏禾试验组释放率一直较高,试验结束时仍然在0.07 d-1左右.植物生长会带来底质氮释放的增加,尤其在试验初期,各试验组释放率要高于空白组.但是随着试验的进行,双穗雀稗和黄菖蒲试验组的释放率都达到空白对照组的水平,这两种植物的抑制氮释放的效果明显.

|
图 3 不同试验组底质对上覆水总氮释放速率 Fig. 3 Release rate of nitrogen from sediment to overlaying water in experimental groups |
如图 4所示,空白对照组沙壤底质总氮含量在试验后下降,修复底质总氮含量试验后小幅上升至650 mg·kg-1;而各试验组的变化情况则相反,各试验组沙壤底质总氮含量上升,修复底质总氮含量维持在600 mg·kg-1.研究空白组与试验组、沙壤底质与修复底质的差异后,可以看出植物再试验期间增加了底质中氮的含量,增量在沙壤底质中尤为明显.相反当底质没有植物的作用时,氮含量则会减少.结合2.1.2节中底质向上覆水氮的释放速率在试验期间普遍高于空白组,可以认为植物在生长过程中,根系分泌的激素、植物蛋白,增加了底质氮的赋存,尤其在颗粒较大的沙壤中,植物的分泌活动愈加旺盛,促进了底质氮的积累.

|
EG1、EG2、EG3、EG4、EG5分别代表双穗雀稗、李氏禾、香菇草、黄花水龙、黄菖蒲试验组,Origin表示试验前底质,Blank表示空白对照组 图 4 试验前后各试验组和空白组底质氮含量 Fig. 4 Nitrogen in sediment of experimental groups at the beginning and the end of test cycle |
如图 5(a)所示,空白试验组上覆水溶解性总磷DTP在试验0~7 d由0.01 mg·L-1增加至0.055 mg·L-1、溶解性无机磷DIP浓度由0.005 mg·L-1增加至0.03 mg·L-1,14 d后,DTP浓度略有上升,DIP浓度则下降至0.02 mg·L-1.由图 5(b)~5(f)可知,试验组DTP和DIP浓度低于空白组,试验开始时各组DTP浓度0.01 mg·L-1,0~7 d各组浓度有不同程度的增长,随后7~20 d DTP维持在一定浓度,DIP则持续下降.空白试验组上覆水DTP、DIP浓度变化情况说明底质中可溶态磷快速地向上覆水释放,在水体中到达一定浓度后在底质-上覆水界面形成平衡状态[15];而各试验组DTP、DIP浓度的普遍偏低和下降说明了植物可以有效地抑制磷的释放.同时可以发现14 d停止扰动是DTP浓度变化的转折点,14 d前DTP浓度快速上升,14 d停止扰动时DTP浓度大幅下降,当第14 d恢复扰动后DTP浓度则持续回升.扰动带来了上覆水DTP浓度的增加,而植物可以在扰动条件下减少释放的量.空白组与各试验组DIP/DTP比例变化的多样性反映了上覆水磷形态受到水体理化条件、微生物、底质磷形态的多重影响[16],扰动不是上覆水磷形态的决定因素.

|
图 5 各组释放过程中上覆水DTP、DIP的浓度及比例变化情况 Fig. 5 Concentration and percentage of different forms of phosphorus (DTP, DIP) in the process of release |
与总氮释放情况相似的是,底质向上覆水总磷释放差异主要在各试验组间,试验组内两种底质的总磷释放没有显著性差异(α < 0.05).如图 6所示空白对照组和各试验组的底质释放TP速率在试验开始时0~7 d最高,空白组达到0.58 d-1,李氏禾、黄花水龙试验组达到0.3 d-1,双穗雀稗、香菇草、黄菖蒲试验组达到0.15 d-1;试验至20 d空白组释放速率下降至0.25 d-1,李氏禾、黄花水龙试验组下降至0.1 d-1,双穗雀稗、香菇草、黄菖蒲试验组下降至0.05 d-1.可见,与空白试验组相比,不同水生植物对底质的TP释放表现出显著的抑制作用,在试验前期就可以快速固定下底质中TP,减少释放速率,在试验过程中能稳定控制释放速率,减少释放总量,使各试验组上覆水TP浓度低于空白对照组.

|
图 6 不同试验组底质对上覆水总磷释放速率 Fig. 6 Release rate of Phosphorus from sediment to overlaying water in experimental groups |
如图 7所示,试验后空白试验组底质中总磷的含量没有明显变化,而有效磷的比例升高;各试验底质总磷、有效磷则有均大幅下降.其中李氏禾、黄花水龙、黄菖蒲试验组底质中TP含量低于100 mg·kg-1, 双穗雀稗试验组下降至300 mg·kg-1,香菇草下降至380 mg·kg-1,各组中有效磷均低于100 mg·kg-1. 2.2.2节中就已经指出了磷的释放得到了抑制,说明植物可以在试验周期内吸收利用底质中的磷.

|
EG1、EG2、EG3、EG4、EG5分别代表双穗雀稗、李氏禾、香菇草、黄花水龙、黄菖蒲试验组,Origin表示试验前底质,Blank表示空白对照组 图 7 试验前后各试验组和空白组底质磷含量 Fig. 7 Phosphorus in sediment of experimental groups at the beginning and the end of test cycle |
图 8中曲线表示小于等于某个粒径占总颗粒物的比例,分布曲线指出试验前后两种底质的粒径分布变化根据所在试验组存在差异.其中修复底质在试验前粒径以≤100 μm为主,占总体的95%;试验后空白对照组基本保持原有分布,各试验组粒径≤50 μm的比例提高,尤其20 μm以下的细小颗粒为主要增长部分.沙壤底质试验前≤50 μm的颗粒虽然超过80%,但50~100 μm的比例较低,≥100 μm的中等颗粒比例较高;图 8中试验后各组的分布曲线表明沙壤底质在部分植物的影响下粒径50~100 μm的颗粒增加,≥100 μm的中等颗粒比例显著下降,粒径≤50 μm的比例可以达到修复底质的水平. 图 9反映的就是沙壤底质在不同植物的作用下粒径分布的变化情况,其中显示了底质颗粒粒径大小以及所占的比例.试验前各组的沙壤底质100~150 μm的中等颗粒比例高,试验后双穗雀稗、李氏禾试验组100~150 μm中等颗粒比例下降,粒径≤50 μm的比例超过95%;试验后黄菖蒲、香菇草试验组粒径≥100 μm比例略有下降,≤50 μm的比例上升至90%;试验后空白对照组、黄花水龙试验组粒径≥100 μm比例小幅降低,总体上与试验前差异微小.综合上述各组具体数据,可以得出随着植物生长,底质颗粒结构发生了变化,粒径≤50 μm颗粒比例升高,颗粒愈发细密.有研究指出水陆交错带的植物根系通过自身须根的穿插挤压以及分泌物显著改变底质理化特征[17].试验前后沙壤底质粒径分布变化证实了双穗雀稗、李氏禾、黄菖蒲可以对底质加以改性,黄花水龙、香菇草对底质粒径的影响则较小.

|
图 8 试验前后粒径分布变化 Fig. 8 Distribution of particle diameter of experimental groups before and after the test |

|
图 9 试验前后沙壤底质粒径构成情况 Fig. 9 Distribution of particle diameter of experimental groups before and after the test |
上覆水中悬浮颗粒物的主要来源是水体扰动过程中上扬的底质颗粒,促进了底质与上覆水之间的物质交换[18].各试验组与空白组扰动持续7 d,上覆水悬浮颗粒物浓度SS达到最高值,其中空白组的增幅大于试验组. 14 d停止扰动,即14(S),各试验组与空白对照组上覆水SS下降至150 mg·L-1左右. 14 d停止扰动6 h后恢复水力扰动后,SS小幅上升,随着试验进行(15~20 d),各试验组中新生底质上覆水SS基本维持在120 mg·L-1.各试验组中沙壤底质上覆水SS存在较大差异(表 2):空白试验组沙壤底质SS超过了220 mg·L-1,黄花水龙在各试验组中最高,达到125 mg·L-1;黄菖蒲最低,接近25 mg·L-1;双穗雀稗、李氏禾、香菇草组均达到100 mg·L-1.对比不同试验组扰动期间的SS变化情况,可以发现黄菖蒲 < 李氏禾 < 香菇草 < 双穗雀稗 < 黄花水龙 < 空白对照组.植物生长起到了抑制底质悬浮,减少悬浮颗粒物浓度的作用.
|
|
表 2 各试验组上覆水悬浮颗粒物浓度/mg·L-1 Table 2 Concentration of Suspended Solid in overlaying water of experiment groups/mg·L-1 |
2.3.3 植物对氮磷释放影响
如图 10所示,5种植物在试验过程中长势良好,试验后生物量增加.黄花水龙是浮叶植物,试验后植物鲜重达到1.2 kg·m-2.李氏禾与双穗雀稗均属于禾本科植物,两者都有着发达的匍匐茎和根壮茎,试验后李氏禾鲜重增加到0.85 kg·m-2, 双穗雀稗鲜重增加到0.65 kg·m-2.黄菖蒲是湿生/挺水植物,枝叶茂盛,根茎短粗,环境适应性较好,鲜重在试验前后有较快的增长,增加至0.75 kg·m-2.香菇草是一种低矮(株高15 cm)的蔓生挺水/湿生植物,生物量远远小于其他较为高大的挺水/湿生植物,试验前后生物量增加至0.52 kg·m-2.

|
图 10 不同植物组试验前后生物量变化 Fig. 10 Biomass of experimental groups before and after the test |
已有研究认为根长密度、根表面积密度、根重密度是反映根系特征的重要指标,植物根系与底质理化性质变化息息相关[19, 20].须根长度密度可以反映一种植物根部须根的伸展情况,须根表面积密度可以反映植物根部和底质的接触情况,根重密度则直接体现须根的数量. 表 3显示在选取的5种水陆交错带植物中,黄菖蒲的d≤1 mm须根根长密度、根表面积密度较高,而李氏禾根重密度较高,5种植物的根系参数有明显的差异,但香菇草与黄菖蒲一样有很强的适应性,可以在试验装置中快速扎根稳定.通过分析须根参数与各影响因素的相关关系,可以得出须根参数与上覆水中氮磷的释放量、底质中氮磷赋存量之间有较强的相关关系(P < 0.05).根据表 4中的相关系数,须根参数与底质磷的赋存量以及向上覆水的释放量均呈负相关,与底质氮的赋存量呈正相关,对上覆水氮的释放量则没有显著影响.
|
|
表 3 5种水陆交错带植物d≤1mm须根特征参数 Table 3 Characteristic parameters of the fine root (d≤1mm) of 5 littoral zone plants |
|
|
表 4 氮磷释放量与影响因素相关矩阵 Table 4 Correlation matrix of N and P release and influential factors |
研究上述植物、底质、上覆水指标数据之间的相关关系发现,底质的颗粒和氮磷赋存情况受到d≤1 mm的须根的直接影响[21],根长密度、根表面积密度、根重密度与粒径小于50 μm的底质颗粒比例有显著的正相关关系(P < 0.05).鲁素云[22]指出部分水生植物根系通过穿插、分泌糖类、有机酸甚至腐败分解等方式促进底质在水体冲击下的稳定.植物根系不仅通过分泌活性酶、植物激素来包裹固结底质,其自身生长对底质的穿插压实让底质的颗粒越来越细密[23].试验显示上覆水SS与≤50 μm的底质颗粒比例呈负相关关系(-0.546*),各试验组中底质颗粒改性能力强弱顺序为双穗雀稗 > 李氏禾 > 黄菖蒲 > 黄花水龙 > 香菇草 > 空白对照组,上覆水SS高低顺序为黄菖蒲 < 李氏禾 < 香菇草 < 双穗雀稗 < 黄花水龙 < 空白对照组,这是植物须根在底质中的生长影响着底质的颗粒粒径,同时须根通过分泌有机物质包裹粘连底质细小颗粒,对底质的穿插挤压也使底质颗粒物质变得更加细小,更易与须根紧密粘连;同时上覆水中SS对氮、磷释放量具有显著正线性关系,是底质氮磷释放的重要途径.
在影响底质氮磷释放的诸多因素中,以植物根长密度、根重密度、根表面积密度3项须根参数为主形成了主要因素1(PAC1),以底质氮磷赋存量形成了主要因素2(PAC2),以植物生物量形成了主要因素3(PAC3).在影响底质氮磷释放的因素中,PAC1和PAC2产生了明显的影响,PAC1显示TP释放量、底质总磷、上覆水SS受植物根长密度、根重密度、根表面积密度的抑制,相关分析也显示出它们之间有显著的负相关关系;PAC2指出底质中TP含量直接影响着对上覆水TP的释放,相关分析显示底质中TP含量越高,TP释放量约高,而底质中TN含量对上覆水TN的释放影响有限,相关性分析也显示两者之间没有显著的相关性. PAC3作为唯一以单一指标形成的主要因素,其对底质理化特征及植物须根的影响较小,但对TN、TP释放量也具有一定影响,相关性分析也显示出植物生物量与TN、TP释放量分别有-0.172*、0.274*显著线性关系(图 11).综上植物对底质氮磷释放有不同的抑制效果,这种抑制作用可以归纳为通过植物根系改变底质的颗粒构成,颗粒构成的改变减少了底质的上扬,间接减少了氮磷的释放量,还通过对底质氮磷赋存量的改变直接影响氮磷的释放;另外,植物根系对磷的吸收利用要远远高于氮,减少了磷尤其是无机态磷的释放;植物根系生长促使底质氮的积累,底质含氮物质在向上覆水释放的过程也比较缓慢,部分植物还会促使上覆水中NH4+-N和NO3--N的比例提高.

|
图 11 底质氮磷释放主因素分析结果 Fig. 11 Main factors affecting the release of nitrogen and phosphorus in sediments |
(1) 植物生长促使底质颗粒结构发生了变化,粒径≤50μm颗粒比例升高,颗粒愈发细密.各试验组中底质颗粒改性能力强弱顺序为双穗雀稗 > 李氏禾 > 黄菖蒲 > 黄花水龙 > 香菇草 > 空白对照组,上覆水SS与底质颗粒构成密切相关,上覆水SS高低顺序为黄菖蒲 < 李氏禾 < 香菇草 < 双穗雀稗 < 黄花水龙 < 空白对照组.
(2) 植物生长增加了底质氮的赋存,尤其在颗粒较大的沙壤中,植物的分泌活动愈加旺盛,促进了底质氮的积累,带来底质氮释放的增加.水体扰动加剧底质向上覆水TN、NH4+-N、NO3--N的释放,但双穗雀稗和黄菖蒲这两种植物有明显的氮释放抑制作用,将释放率控制在0.02 d-1,上覆水TN、NH4+-N、NO3--N浓度控制在0.4、0.2、0.05 mg·L-1.
(3) 植物对底质的TP释放表现出显著的抑制作用,在试验前期就可以快速固定下底质中TP,减少释放速率,双穗雀稗、香菇草、黄菖蒲可以将上覆水TP浓度控制在0.02 mg·L-1,而李氏禾、黄花水龙则为0.03 mg·L-1;李氏禾、黄花水龙试验组TP释放率控制在0.1 d-1,双穗雀稗、香菇草、黄菖蒲试验组控制在0.05 d-1.
(4) 植物对底质氮磷释放有不同的抑制效果,这种抑制作用可以归纳为通过植物根系改变底质的颗粒构成,颗粒构成的改变减少了底质的上扬,间接减少了氮磷的释放量,还通过对底质氮磷赋存量的改变直接影响氮磷的释放;另外,植物根系对磷的吸收利用要远远高于氮,减少了磷尤其是无机态磷的释放;植物根系生长促使底质氮的积累,底质含氮物质在向上覆水释放的过程也比较缓慢,部分植物还会使NH4+-N、N03--N的比例提高.几种水陆交错带植物正是通过上述机制抑制底质中氮磷的释放,其中黄菖蒲、双穗雀稗因为其发达的根系,抑制氮磷物质释放的效果最佳,其次为李氏禾、香菇草,黄花水龙对底质氮磷释放的抑制作用在5种植物中最小.
| [1] | 赵晓峰.太湖湖滨带水生植物时空分布特征及其与水质因子的相互关系研究[D].北京:中国环境科学研究院, 2012. Zhao X F. The study on relationship between the spacial and temporal distribution characteristic of aquatic macrophyte and water quality factors in the Littoral Zone of Taihu Lake[D]. Beijing:Chinese Research Academy of Environmental Sciences, 2012. http://cn.bing.com/academic/profile?id=86a2cd9ca07eb5445bf41f8b476bdd8a&encoded=0&v=paper_preview&mkt=zh-cn |
| [2] | 谢杰, 吴德意, 陈雪初, 等. 滇池和洱海湖滨带水生植被状况与水质的关系研究[J]. 环境科学与技术, 2013, 36(2) : 55–59. Xie J, Wu D Y, Chen X C, et al. Relationship between aquatic vegetation and water quality in littoral zones of Lake Dianchi and Lake Erhai[J]. Environmental Science & Technology, 2013, 36(2) : 55–59. |
| [3] | 方焰星, 何池全, 梁霞, 等. 水生植物对污染水体氮磷的净化效果研究[J]. 水生态学杂志, 2010, 3(6) : 36–40. Fang Y X, He C Q, Liang X, et al. The purifying effect of polluted water by the aquatic plants[J]. Journal of Hydroecology, 2010, 3(6) : 36–40. |
| [4] | 甘树, 卢少勇, 秦普丰, 等. 太湖西岸湖滨带沉积物氮磷有机质分布及评价[J]. 环境科学, 2012, 33(9) : 3064–3069. Gan S, Lu S Y, Qin P F, et al. Spatial distribution and evaluation of nitrogen, phosphorus and organic matter in surface sediments from western lakeside belt of Lake Taihu[J]. Environmental Science, 2012, 33(9) : 3064–3069. |
| [5] | 尹延震, 储昭升, 赵明, 等. 洱海湖滨带水质的时空变化规律[J]. 中国环境科学, 2011, 31(7) : 1192–1196. Yin Y Z, Chu Z S, Zhao M, et al. Spatial and temporal changes in water quality in aquatic-terrestrial ecotone of Lake Erhai[J]. China Environmental Science, 2011, 31(7) : 1192–1196. |
| [6] | 叶春.退化湖滨带水生植物恢复技术及工程示范研究[D].上海:上海交通大学, 2007. Ye C. Studies on macrophyte restoration in the damaged aquatic-terrestrial ecotone and engineering demonstration[D]. Shanghai:Shanghai Jiao Tong University, 2007. http://www.oalib.com/references/15703264 |
| [7] | 王磊, 李冬林, 丁晶晶, 等. 太湖湖滨湿地沉积物氮磷与2种挺水植物氮磷的关系[J]. 生态环境学报, 2011, 20(10) : 1523–1529. Wang L, Li D L, Ding J J, et al. Relationship between N and P contents in sediments and two emerged plants in Taihu Lakeside wetland[J]. Ecology and Environmental Sciences, 2011, 20(10) : 1523–1529. |
| [8] | Horppila J, Nurminen L. Effects of submerged macrophytes on sediment resuspension and internal phosphorus loading in Lake Hiidenvesi (southern Finland)[J]. Water Research, 2003, 37(18) : 4468–4474. DOI: 10.1016/S0043-1354(03)00405-6 |
| [9] | Özbay H. Changes in water properties in two different macrophyte stands and in open water in a shallow freshwater lake in Turkey[J]. Fresenius Environmental Bulletin, 2014, 23(5) : 1150–1154. |
| [10] | 伍清新, 刘杰, 游少鸿, 等. 李氏禾湿地系统净化Cr (Ⅵ)污染水体的机理研究[J]. 环境科学学报, 2014, 34(9) : 2306–2312. Wu Q X, Liu J, You S H, et al. Decontamination mechanism of Cr (Ⅵ)-polluted water in constructed wetland planted with Leersia hexandra Swartz[J]. Acta Scientiae Circumstantiae, 2014, 34(9) : 2306–2312. |
| [11] | 刘丹, 白画画, 朱元荣, 等. 太湖典型植物氨基酸组成特征及其对水环境的影响[J]. 湖泊科学, 2015, 27(1) : 119–127. Liu D, Bai H H, Zhu Y R, et al. Characteristics and influence to overlying water of amino acids from typical plants distributed in Lake Taihu, China[J]. Journal of Lake Sciences, 2015, 27(1) : 119–127. DOI: 10.18307/2015.0115 |
| [12] | 赵凯, 李振国, 魏宏农, 等. 太湖贡湖湾水生植被分布现状(2012年)[J]. 湖泊科学, 2015, 27(3) : 421–428. Zhao K, Li Z G, Wei H N, et al. The distribution of aquatic vegetation in Gonghu Bay, Lake Taihu, 2012[J]. Journal of Lake Sciences, 2015, 27(3) : 421–428. DOI: 10.18307/2015.0308 |
| [13] | 向速林, 朱梦圆, 朱广伟, 等. 太湖东部湖湾大型水生植物分布对水质的影响[J]. 中国环境科学, 2014, 34(11) : 2881–2887. Xiang S L, Zhu M Y, Zheng G W, et al. Influence of macrophytes on water quality in the eastern bays of Lake Taihu, China[J]. China Environmental Science, 2014, 34(11) : 2881–2887. |
| [14] | 徐新洲, 薛建辉, 吕志刚, 等. 太湖贡湖湾湖滨湿地生态功能区与植被修复研究[J]. 南京林业大学学报(自然科学版), 2013, 37(3) : 35–40. Xu X Z, Xue J H, Lv Z G, et al. A research of ecological function area and vegetation restoration at Taihu Gonghu bay wetland[J]. Journal of Nanjing Forestry University (Natural Sciences Edition), 2013, 37(3) : 35–40. |
| [15] | 何东, 张毅敏, 杨飞, 等. 太湖藻源性颗粒物降解过程中营养盐转化及其生态效应[J]. 中国环境科学, 2016, 36(3) : 899–907. He D, Zhang Y M, Yang F, et al. The transformation of the nutrient in the degradation process of the phytoplankton-derived particulate organic matter and it's ecological effect[J]. China Environment Science, 2016, 36(3) : 899–907. |
| [16] | 何佳, 陈春瑜, 邓伟明, 等. 滇池水-沉积物界面磷形态分布及潜在释放特征[J]. 湖泊科学, 2015, 27(5) : 799–810. He J, Chen C Y, Deng W M, et al. Distribution and release characteristics of phosphorus in water-sediment interface of Lake Dianchi[J]. Journal of Lake Sciences, 2015, 27(5) : 799–810. DOI: 10.18307/2015.0506 |
| [17] | 徐少君, 曾波, 类淑桐, 等. 三峡库区几种耐水淹植物根系特征与土壤抗水蚀增强效应[J]. 土壤学报, 2011, 48(1) : 160–167. Xu S J, Zeng B, Lei S T, et al. Root features of several flooding-tolerant plants and their roles in enhancing anti-erodibility of the soil in three gorges reservoir region[J]. Acta Pedologica Sinica, 2011, 48(1) : 160–167. |
| [18] | 郭俊锐, 李大鹏, 刘焱见. 扰动对悬浮颗粒物粒径及上覆水中磷形态分布的影响[J]. 环境科学, 2016, 37(4) : 1422–1426. Guo J R, Li D P, Liu Y J. Impacts of sediment disturbance on the distribution of suspended particle size and phosphorus[J]. Environmental Science, 2016, 37(4) : 1422–1426. |
| [19] | 卢少勇, 曲洁婷, 许秋瑾, 等. 贡湖湾北部退渔还湖区水、沉积物和土壤中氮的时空分布[J]. 农业环境科学学报, 2014, 33(11) : 2234–2241. Lu S Y, Qu J T, Xu Q J, et al. Distribution of nitrogen in water, soil and sediment in water/land ecotone of returning fishpond to lake area of Northern Gonghu Bay[J]. Journal of Agro-Environment Science, 2014, 33(11) : 2234–2241. |
| [20] | Janadeleh H, Alhashemi A H, Nabavi S M B. Investigation on concentration of elements in wetland sediments and aquatic plants[J]. Global Journal of Environmental Science and Management, 2016, 2(1) : 87–93. |
| [21] | 丁庆章, 刘学勤, 张晓可. 水位波动对长江中下游湖泊湖滨带底质环境的影响[J]. 湖泊科学, 2014, 26(3) : 340–348. Ding Q Z, Liu X Q, Zhang X K. Impacts of water level fluctuations on substrate environments of lakeshore zone of the lakes in the middle and lower reaches of the Yangtze River[J]. Journal of Lake Sciences, 2014, 26(3) : 340–348. DOI: 10.18307/2014.0302 |
| [22] | 鲁素芸. 植物根际生态学与根病生物防治进展[M]. 北京: 中国人民大学出版社, 1990. |
| [23] | 李强.黄土丘陵区植物根系强化土壤抗冲性机理及固土效应[D].北京:中国科学院教育部水土保持与生态环境研究中心, 2014. Li Q. Mechanism of plant roots in improving resistance of soil to concentrated flow erosion and reinforcement in loess hilly region[D]. Beijing:Research Center of Soil and Water Conservation and Ecological Environment, Chinese Academy of Sciences and Ministry of Education, 2014. http://cn.bing.com/academic/profile?id=654640633c584f0e4f6cf7554c2130b3&encoded=0&v=paper_preview&mkt=zh-cn |