 2017, Vol. 38
2017, Vol. 38


2. 华东师范大学河口海岸学国家重点实验室, 上海 200062
2. State Key Laboratory of Estuarine and Coastal Research, East China Normal University, Shanghai 200062, China
湖泊富营养化导致的藻类暴发一直是我国最为突出的水环境问题之一[1]. 藻类过度生长是多种因素共同作用的结果,既包括水温、 光照、 风速等自然因素,也包括氮(N)、 磷(P)、 铁(Fe)等营养物过量排放的人类活动因素[2, 3]. 在诸多因素中,全球气候变暖背景下的水温变化与高强度人类活动所引起的N、 P排放增加被认为是导致湖泊富营养化最关键因素,因此,同时考虑水温、 N、 P因子变化的湖泊富营养化相关研究在逐渐增多[4~6],但温度与营养物对湖泊藻类生物量的交互作用等还需要深入研究[7],比如水温、 N、 P促进藻类生物量增长的相对重要度的长期变化规律和季节性特征.
富营养化湖泊的藻类生长是自然界中一个非期望或非平均的现象,藻类生物量数据异质性很强,水华期间的藻类数据会呈“高峰厚尾”的分布,或存在显著的异方差等情况[8]. 近年来在环境科学和生态学领域受到重视的分位数回归(quantile regression)方法特别适合处理这种波动性大、 异质性很强的环境数据[9, 10]. 该方法可针对回归变量任何一个分位点进行回归分析,且在存在极端值或重尾情况时仍能保持较好的稳健性,适宜处理应变量对自变量的极端响应,而不只是平均水平的响应[11~13],因此能更加全面地反映藻类生物量对水温、 N、 P等环境指标的响应特征. 本研究基于云南洱海长时间尺度(1990-2013年)的水质观测数据,运用分位数回归方法,按不同年份区间和不同季节分别分析洱海藻类生物量[以叶绿素a(Chl-a)表征]对N、 P、 水温的定量响应关系,探讨营养物因子与水温因子相对重要性的长时间尺度演变规律和季节性变化规律,对制定洱海富营养化控制策略提供科学依据.
1 材料与方法 1.1 研究区域洱海是云南省第二大高原淡水湖泊,为滇西最大的断陷湖,跨洱源、 大理两县市,处于东经100°06′-100°17′,北纬25°36′-25°55′之间. 水面面积249.80 km2,汇水面积2 565.0 km2,最大水深21.0 m,平均水深10.5 m,库容28.8亿m3(图 1). 洱海是沿湖人民生活、 灌溉、 工业用水的主要水源地,是整个流域社会经济可持续发展的基础[14]. 洱海湖面多年平均海拔1 965.8 m,光照充足,辐射强,气温温和,为浮游藻类的大量繁殖提供了有利条件. 区年均气温15℃左右,年均降雨量1 055 mm,年均蒸发量1 970 mm. 流域水系发达,入湖河流大小共 117条.

|
图 1 洱海流域及水质监测点分布示意 Fig. 1 Lake Erhai watershed and water sampling sites |
从云南省环境监测中心站、 中国大理洱海湖泊研究中心等环境监测和研究部门收集了洱海1990-2013年的水环境常规监测数据,全湖水质监测点为13个(图 1),经纬度依次为: 25°51′36″N 100°10′12″E,25°51′18″N 100°11′24″ E,25°51′00″N 100°12′36″E,25°41′42″N 100°12′54″E,25°42′00″N 100°13′48″E,25°42′18″N 100°15′00″E,25°37′48″N 100°13′48″E,25°36′0″N 100°15′0″E,25°37′60″N 100°15′50″E,25°55′30″N 100° 6′54″E,25°54′54″N 100°8′42″E,25°54′36″N 100°10′48″E,25°47′29″N 100°11′43″E. 水质指标主要包括总氮(TN)、 总磷(TP)、 水温、 藻类叶绿素a(Chl-a),采样频次为一年6次或12次. 各水质指标的主要测试方法如下:TN为过硫酸钾氧化紫外分光光度法,TP为钼锑抗分光光度法,水温为温度计现场实时测定,Chl-a为四波段分光光度法[15]. 数据主要统计特征见表 1,TP的变异系数最大(239.7%),其次是Chl-a和TN,这可能与各指标的年份间差异和季节性差异有关. 各指标数据的Kolmogorov-Smirnov正态分布检验,除了水温数据呈正态分布以外,所有指标的数据都呈非正态分布.
|
|
表 1 洱海水温、 总氮、 总磷及藻类叶绿素a的统计值 (n=1 419) Table 1 Descriptive statistics and normality test of water temperature,TN,TP and algal Chl-a of Lake Erhai recorded between 1991 and 2011 |
1.3 分位数回归方法
分位数回归模型(quantile regression)是依据因变量的条件分位数对自变量进行回归,得到所有分位水平的回归模型[16, 17].
假设随机变量的分布函数为F(y)=Prob(Y≤y),Y 的τ 分位数为满足F(y) ≥τ的最小y 值:
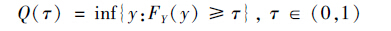
|
F(y)的τ 分位点Q(τ)由最小化关于ξ 的目标函数得到:
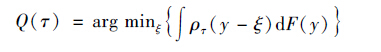
|
其中ρτ(u)称为检验函数:

|
假设因变量Y和自变量X 在τ 分位的线性函数关系为Y=X′β+ε,给定X=x时,Y的条件分布函数为FY (y | x),则τ分位数为:
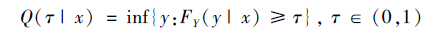
|
线性条件分位数通常表示为:
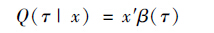
|
分位数回归能在不同的分位数τ得到不同的分位函数,残差计算方法不同于最小二乘法,具体如下:
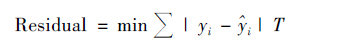
|
考虑长时间尺度变化,将1990-2013年的监测数据按照相同年限(6 a)分成4组:1990-1995年、 1996-2001年、 2002-2007年、 2008-2013年,分别对这4组进行三元分位数回归分析. 为分析季节变化,将所有监测数据按春、 夏、 秋、 冬四季分成4组,按不同季节分别进行三元分位数回归分析,比较各因子的斜率值变化,并计算95%置信区间(CI),CI覆盖0(包含0),表示相关性“不显著”,而CI未覆盖0(不包含0),说明相关性“显著”. 为便于比较3个因子的斜率值变化,在进行多元分位数回归之前,所有原始数据取对数(lg) 处理.
使用统计软件STATA V.12.0进行分位数回归分析和参数估计.
2 结果与讨论 2.1 水质指标的年际变化与季节变化 2.1.1 年际变化从TN变化趋势图看,2001年之前,TN浓度呈缓慢上升趋势,但总体处于较好的Ⅱ类水质. 在2001-2003年期间有一次跳跃式增长,从Ⅱ类区跨入Ⅲ类区,近10年(2003-2013年)TN浓度维持在Ⅲ类水平,达到2001年之前浓度水平的近2倍[图 2(a)]. TP浓度年均值的变化趋势表现为有升有降,基本在Ⅱ类水平线上下浮动,浓度高峰出现在2002-2004年区间和2013年. 近10余年(2002-2013年)的TP浓度水平比前10年(1990-2001年)的浓度水平总体高出15%左右[图 2(b)]. 洱海的年平均水温在16-19℃之间周期性波动,未见明显的上升或下降趋势[图 2(c)].

|
图 2 洱海TN、 TP、 水温和藻类Chl-a的逐年变化 (1990-2013年) Fig. 2 Yearly variation of TN,TP,water temperature,algal Chl-a in Lake Erhai from 1990 to 2013 |
从藻类Chl-a变化趋势看,2000年之前洱海全湖Chl-a维持在低浓度水平,变化范围为1.0-3.5 mg·m-3. 2000-2002的3年期间,Chl-a浓度呈直线上升,藻类生物量从低水平跨越至较高水平. 2003年之后的Chl-a平均浓度达到2000年之前的10余倍[图 2(d)]. Pearson相关性分析显示藻类Chl-a与TN以及水温均呈显著正相关性(P<0.01),而与TP的相关性不显著(P>0.05).
2.1.2 季节变化4项水质指标具有明显的季节性特征,均表现为夏、 秋两季较高,且两季数据接近,而春、 冬季数值较低. 8-10月TN浓度最高,5月是一年中TN上升的起点,11月是TN开始下降的拐点[图 3(a)]. 一年中TP浓度在4-7月是洱海上升速率最快的时段[图 3(b)]. 水温在夏季和秋季最高,分别为23.3℃和20.2℃,很适宜藻类生长[图 3(c)]. 藻类Chl-a浓度在6-7月出现跳跃式上升,平均值由10 mg·m-3跃升至20 mg·m-3,在7-11月维持在最高水平[图 3(d)].

|
图 3 洱海TN、 TP、 水温和藻类Chl-a的季节变化 Fig. 3 Seasonal variation of TN,TP,water temperature,algal Chl-a in Lake Erhai |
通过三元分位数回归方法分析洱海N、 P和水温对藻类Chl-a的耦合效应随时间变化特征. 4个年份区间的lg(TN)、 lg(TP)和lg(水温)的斜率值随分位点的变化谱图见图 4,斜率值及其95%置信区间的统计特征见表 2.
|
|
表 2 不同年份区间的多元分位数回归的斜率参数统计 Table 2 Statistics of all estimated parameters obtained by multivariate QR and OLSR for the three lakes in different periods |

|
图 4 4个年份区间的三元分位数回归中lg(TN)、 lg(TP)和lg(Temp)的斜率变化 Fig. 4 Estimated slopes based on triple quantile regression of lg(Chl-a) vs. lg(TN) & lg(TP) & lg(Temp) in the four periods |
4个时间段lg(TN)、 lg(TP)和lg(水温)的平均斜率值变化范围分别为0.03-0.20、 -0.30-0.80、 0.44-2.70,3个因子对藻类的促进效应依次为水温>磷>氮. 各因子的斜率值随时间变化很大,水温的斜率值逐渐下降,从1990-1995年期间的2.05-4.78快速下降至2008-2013期间的0.15-0.75,说明水温对藻类生长的正效应在持续相对下降. 水温斜率的95%置信区间(CI)在2007年之前的时间段全未覆盖0,而在2008-2013年期间只有部分区间(0.45<τ<0.71)未覆盖0,说明水温与藻类Chl-a之间相关性总体表现为极显著,但在2008年之后显著度出现下降. 相反,TP的斜率值逐渐上升,从1990-1995年期间的负值(-0.42--0.05)快速攀升至2008-2013年期间的0.52-1.07,说明P对藻类生长的正效应逐渐在相对增强. 除了1996-2001年时间段,TP斜率的95%CI在绝大部分位点均是未覆盖0,说明TP与藻类Chl-a之间相关性总体呈极显著. 在过去20余年中TN的斜率值比较低,但在绝大部分分位条件下表现为正效应,在1996-2001年和2002-2007年两个时段的数值略高,而在2008-2013年的正效应达到最低. TN斜率的95%CI只在1996-2007年期间的部分分位点未覆盖0,说明TN与藻类Chl-a之间相关显著度较低.
总体上看,水温和营养物共同支撑着洱海藻类生物量,这与国内外其他富营养化湖泊的研究结果类似[18, 19]. 营养物和水温对藻类生物量变化的耦合效应比较复杂,据1980-2014年的洱海陆域大理站气象资料,当地日平均气温、 日最高气温以及日最低气温均存在缓慢上升的趋势,近30余年日最低气温上升幅度为0.05℃·a-1,有气候变暖趋势[20]. 而分位数回归结果显示水温是洱海藻类增长的重要限制因素,但其相对重要度随着时间推移(富营养化程度加重)明显下降,而P的相对重要度明显持续上升,P比N对藻类的限制作用更强. 与水温相比,营养盐浓度可能是影响洱海藻类生物量的更重要因素. 这与Jeppesen等[21]提出的亚热带到温带区域的35个湖泊中浮游藻类生物量与结构主要是由营养物决定,而气候变暖的作用较小的结论基本一致.
分位数回归分析结果显示洱海藻类属于N、 P共同限制,这与N/P比值有关. 一般当N/P质量比在10∶1-25∶1的范围时,藻类生长出现氮磷共同限制的概率大[22~24]. 洱海N/P质量比值一般在10-30之间波动,平均值为23,中位数21(表 1),符合藻类出现N、 P共同限制的条件. 藻类Chl-a对营养物和水温的响应关系可能与藻类群落结构演替有很大关系. 洱海的富营养化演进过程是藻类群落结构逐渐从硅藻门占优势向蓝藻门占优势的演变过程[25]. 不同藻类对N、 P和水温的响应程度差别会很大,硅藻的最主要限制因子可能是水温等气象条件,其次是营养盐[26],而蓝藻一般受N/P质量比值的影响显著[27, 28],还有水温、 光照、 气压等条件[29]. 洱海N/P质量比值现状适合微囊藻、 鱼腥藻等蓝藻门在藻类竞争中占优势.
2.3 藻类对氮、 磷及水温响应的季节性特征不同季节中lg(TN)、 lg(TP)和lg(水温)的斜率值随分位点的变化谱图见图 5. 春季,TN和水温的斜率值均始终保持为正值,两者的95% CI在绝大部分分位条件下均未覆盖0(极显著). TP斜率在大多数条件下(τ<0.85)为负值. 在较高分位时(τ>0.7,Chl-a>5.4 mg·m-3),水温和TN的斜率值同时下降,而TP斜率值明显相应上升,当τ>0.85处(Chl-a>8.0 mg·m-3)TP的斜率值转为正. 这说明N和水温是支撑春季藻类生物量的重要因素,但出现高生物量的决定性因子是P.

|
图 5 分季节的三元分位数回归中lg(TN)、 lg(TP)和lg(Temp)的斜率值变化 Fig. 5 Estimated slopes based on triple quantile regression of lg(Chl-a) vs. lg(TN) & lg(TP) & lg(Temp) for four seasons |
夏季,水温的斜率值在所有分位点全部为负值,数值范围为-2.27-0.03,而TN和TP斜率平均值分别为0.51和0.26,在较高分位时两者的95% CI均未覆盖0(极显著). 当τ<0.40(Chl-a<3mg·m-3),TN斜率持续上升,而TP斜率相应下降. 而当τ>0.40,TN斜率持续缓慢下降,而TP斜率持续快速上升,说明夏季藻类是N、 P共同限制,但随着藻类生物量增长,TP对藻类的正效应逐渐强于TN.
秋季,TN和水温的斜率全部为正值,平均值分别为0.66和4.59,两者的95% CI在所有分位点都未覆盖0(极显著). TP斜率总体是大于0,平均值0.16,而95%CI在绝大部分分位条件下是覆盖0(不显著). 秋季藻类增长的限制因子重要度排序为:水温>TN>TP.
冬季,TN斜率始终为负值,TP斜率虽然在绝大部分条件下为正值,但其95%CI全覆盖0(不显著). 温度的斜率始终保持正值,其95% CI在绝大部分条件下(0.22<τ<1.52)未覆盖0(极显著),说明水温是冬季藻类增长的主导限制因子.
TN、 TP、 水温、 Chl-a的数值季节性变化表现出高度一致,均是在夏、 秋两季达到最高(图 3),这反映了营养物与水温共同促进洱海藻类生物量变化. 但分位数回归结果显示藻类对各因子的响应关系在夏季和秋季完全不同,夏季日平均气温最高,普遍达到藻类生长所需要的最佳温度,因此水温不属于夏季藻类生长的限制因子. 夏季降雨量大,暴雨径流携带更多营养物进入湖体,促进藻类暴发,N、 P成为藻类共同限制因子[30]. 夏季持续高温还会加速下层水中有机物质的耗氧分解,造成溶解氧浓度急剧下降,容易出现缺氧状态,促进底泥中大量氨氮和磷酸盐加快溶出,致使水中TN、 TP含量升高,支撑全湖藻类出现暴发性增长[31]. 秋季,经过夏季藻类暴发已消耗了大量N、 P,而且秋季气温昼夜温差加大,水温数据的离散度明显大于夏季,藻类生物量对水温变化表现可能更加敏感,使水温成为藻类主要限制因子. 春季和冬季,营养物浓度水平较低,湖水呈相对清洁状态,水温的波动对藻类生物量影响也比较大[32].
洱海藻类群落的季节性特征很明显,在春季以硅藻和蓝藻占优势,冬季以绿藻和蓝藻占优势,而蓝藻在全年绝大部分时间占优势,尤其是在夏秋两季处于绝对优势地位,水华发生时蓝藻数量高达107 cells·L-1[25]. 夏季水温能普遍达到蓝藻生长最理想水温(20℃以上),营养物浓度则成为藻类生物量的决定性因子[33].
3 结论(1) 过去20余年,洱海富营养化程度持续加剧,TN、 TP和Chl-a在2001-2003期间出现跳跃式增长,2003年后的TN平均浓度达到2001年之前浓度水平的近2倍,TP浓度水平比2001年之前的浓度水平总体高出15%,Chl-a平均浓度达到2001年之前的10余倍. 4项水质指标具有明显的季节性特征,均表现为夏、 秋两季较高,且两季数据接近,而春、 冬季数值较低.
(2) 在长时间变化方面,洱海藻类生物量受水温、 N、 P的共同限制作用,但随着洱海富营养化程度的加剧,水温对藻类生物量的相对重要度在持续下降. 营养物对藻类的促进效应在相应提升,尤其是TP对藻类的相对重要度持续攀升. TN对藻类生物量整体表现为较稳定的正效应,但在2008-2013年TN对藻类正效应达到最低. 1995年后,N逐渐取代水温很大部分对藻类的限制作用; 2002之后,P又逐渐取代N成为最重要的营养物限制因子.
(3) 在季节方面,春季和秋季以水温和N的共同限制为主导,但春季当Chl-a>8 mg·m-3时,N和水温对藻类的正效应逐渐被P取代. 夏季完全以N、 P共同限制效应占主导,当Chl-a>3 mg·m-3时,TP的正效应逐渐超过TN,冬季则以P和水温的共同限制效应为主导.
(4) 在气候变暖的大背景下,洱海富营养化控制应采取“控P”为主和N、 P协同削减的策略. 当前富营养化控制过程中,应着重监控春、 秋两季温度短期快速上升可能引发的全湖藻华风险,作好监测与预测工作.
| [1] | 金相灿. 湖泊富营养化研究中的主要科学问题--代"湖泊富营养化研究"专栏序言[J]. 环境科学学报, 2008, 28(1) : 21–23. Jin X C. The key scientific problems in lake eutrophication studies[J]. Acta Scientiae Circumstantiae, 2008, 28(1) : 21–23. |
| [2] | Jung S W, Joo H M, Kim Y O, et al. Effects of temperature and nutrient depletion and reintroduction on growth of Stephanodiscus hantzschii (Bacillariophyceae):implications for the blooming mechanism[J]. Journal of Freshwater Ecology, 2011, 26(1) : 115–121. DOI: 10.1080/02705060.2011.553927 |
| [3] | Symons C C, Arnott S E, Sweetman J N. Nutrient limitation of phytoplankton communities in subarctic lakes and ponds in Wapusk National Park, Canada[J]. Polar Biology, 2012, 35(4) : 481–489. DOI: 10.1007/s00300-011-1092-0 |
| [4] | Fragoso Jr C R, Marques D M L M, Ferreira T F, et al. Potential effects of climate change and eutrophication on a large subtropical shallow lake[J]. Environmental Modelling & Software, 2011, 26(11) : 1337–1348. |
| [5] | 苏洁琼, 王烜, 杨志峰. 考虑气候因子变化的湖泊富营养化模型研究进展[J]. 应用生态学报, 2012, 23(11) : 3197–3206. Su J Q, Wang X, Yang Z F. Lake eutrophication modeling in considering climatic factors change:a review[J]. Chinese Journal of Applied Ecology, 2012, 23(11) : 3197–3206. |
| [6] | 吴功果, 倪乐意, 曹特, 等. 气候变暖和营养水平对洱海浮游植物的长期影响(1980-2009年)[J]. 湖泊科学, 2013, 25(2) : 209–212. Wu G G, Ni L Y, Cao T, et al. Warming and nutrients affect the growth of phytoplankton in Lake Erhai over the period of 1980-2009[J]. Journal of Lake Sciences, 2013, 25(2) : 209–212. DOI: 10.18307/2013.0205 |
| [7] | 董静, 高云霓, 李根保. 淡水湖泊浮游藻类对富营养化和气候变暖的响应[J]. 水生生物学报, 2016, 40(3) : 615–623. Dong J, Gao Y N, Li G B. A review:responses of phytoplankton communities to eutrophication and climate warming in freshwater lakes[J]. Acta Hydrobiologica Sinica, 2016, 40(3) : 615–623. |
| [8] | 李小平, 程曦, 陈小华. 淀山湖营养物输入响应关系的分位数回归分析[J]. 中国环境科学, 2012, 32(2) : 324–329. Li X P, Cheng X, Chen X H. A quantile regression for relationship between nutrient input and response of Dianshan Lake[J]. China Environmental Science, 2012, 32(2) : 324–329. |
| [9] | Planque B, Buffaz L. Quantile regression models for fish recruitment-environment relationships:four case studies[J]. Marine Ecology Progress Series, 2008, 357 : 213–223. DOI: 10.3354/meps07274 |
| [10] | Chen X H, Li X P. Using quantile regression to analyze the stressor-response relationships between nutrient levels and algal biomass in three shallow lakes of the Lake Taihu Basin, China[J]. Chinese Science Bulletin, 2014, 59(28) : 3621–3629. DOI: 10.1007/s11434-014-0553-6 |
| [11] | Friederich P, Hense A. Statistical downscaling of extreme precipitation events using censored quantile regression[J]. Monthly Weather Review, 2007, 135(6) : 2365–2378. DOI: 10.1175/MWR3403.1 |
| [12] | Fleeger J W, Johnson D S, Carman K R, et al. The response of nematodes to deep-sea CO2 sequestration:a quantile regression approach[J]. Deep Sea Research Part Ⅰ:Oceanographic Research Papers, 2010, 57(5) : 696–707. DOI: 10.1016/j.dsr.2010.03.003 |
| [13] | Ebru Ç, Eban A. Determinants of house prices in Istanbul:a quantile regression approach[J]. Quality & Quantity, 2011, 45(2) : 305–317. |
| [14] | 韩涛, 彭文启, 李怀恩, 等. 洱海水体富营养化的演变及其研究进展[J]. 中国水利水电科学研究院学报, 2005, 3(1) : 71–73. Han T, Peng W Q, Li H E, et al. Evolution of eutrophication in the Erhai Lake and its relevant research progress[J]. Journal of China Institute of Water Resources and Hydropower Research, 2005, 3(1) : 71–73. |
| [15] | 国家环境保护总局. 水和废水监测分析方法[M]. (第四版).北京: 中国环境科学出版社, 2002: 50-70. The State Environmental Protection Administration. Water and wastewater monitoring analysis method[M]. (4th ed.). Beijing: China Environmental Science Press, 2002: 50-70. |
| [16] | Cade B S, Terrell J W, Schroeder R L. Estimating effects of limiting factors with regression quantiles[J]. Ecology, 1999, 80(1) : 311–323. DOI: 10.1890/0012-9658(1999)080[0311:EEOLFW]2.0.CO;2 |
| [17] | Cade B S, Noon B R. A gentle introduction to quantile regression for ecologists[J]. Frontiers in Ecology and the Environment, 2003, 1(8) : 412–420. DOI: 10.1890/1540-9295(2003)001[0412:AGITQR]2.0.CO;2 |
| [18] | Paerl H W, Huisman J. Blooms like it hot[J]. Science, 2008, 320(5872) : 57–58. DOI: 10.1126/science.1155398 |
| [19] | Ye C, Shen Z M, Zhang T, et al. Long-term joint effect of nutrients and temperature increase on algal growth in Lake Taihu, China[J]. Journal of Environmental Sciences, 2011, 23(2) : 222–227. DOI: 10.1016/S1001-0742(10)60396-8 |
| [20] | 陈小华. 富营养化初期湖泊(洱海)的环境演变及营养物基准/标准研究[D]. 上海:华东师范大学, 2015. 56-100. Chen X H. Study on the recent 20-year eutrophication process and development of nutrient criteria/standards of Lake Erhai at its initial stage of eutrophication[D]. Shanghai:East China Normal University, 2015. 56-100. |
| [21] | Jeppesen E, Søndergaard M, Jensen J P, et al. Lake responses to reduced nutrient loading-an analysis of contemporary long-term data from 35 case studies[J]. Freshwater Biology, 2005, 50(10) : 1747–1771. DOI: 10.1111/fwb.2005.50.issue-10 |
| [22] | Elser J J, Bracken M E S, Cleland E E, et al. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems[J]. Ecology Letters, 2007, 10(12) : 1135–1142. DOI: 10.1111/ele.2007.10.issue-12 |
| [23] | Wang H J, Liang X M, Jiang P H, et al. TN:TP ratio and planktivorous fish do not affect nutrient-chlorophyll relationships in shallow lakes[J]. Freshwater Biology, 2008, 53(5) : 935–944. DOI: 10.1111/j.1365-2427.2007.01950.x |
| [24] | 陈建良, 胡明明, 周怀东, 等. 洱海蓝藻水华暴发期浮游植物群落变化及影响因素[J]. 水生生物学报, 2015, 39(1) : 24–28. Chen J L, Hu M M, Zhou H D, et al. Studies on population dynamics and the underlying impact factors of phytoplankton during the cyanobacteria bloom in Lake Erhai[J]. Acta Hydrobiologica Sinica, 2015, 39(1) : 24–28. |
| [25] | 胡竹君, 李艳玲, 李嗣新. 洱海硅藻群落结构的时空分布及其与环境因子间的关系[J]. 湖泊科学, 2012, 24(3) : 400–408. Hu Z J, Li Y L, Li S X. Spatial and temporal distributions of diatom communities and their relationships with environmental factors in Lake Erhai[J]. Journal of Lake Sciences, 2012, 24(3) : 400–408. DOI: 10.18307/2012.0311 |
| [26] | 孟顺龙, 裘丽萍, 胡庚东, 等. 氮磷比对两种蓝藻生长及竞争的影响[J]. 农业环境科学学报, 2012, 31(7) : 1438–1444. Meng S L, Qiu L P, Hu G D, et al. Effect of nitrogen and phosphorus ratios on growth and competition of two blue-green algae[J]. Journal of Agro-Environment Science, 2012, 31(7) : 1438–1444. |
| [27] | 王菁, 裘丽萍, 孟顺龙, 等. 氮磷比对鱼腥藻和普通小球藻生长竞争的影响[J]. 江苏农业科学, 2014, 42(5) : 316–320. |
| [28] | 李小平. 蓝藻运动与水华早期预防和控制[J]. 自然杂志, 2008, 30(5) : 280–286. Li X P. Cyanobacteria movements and its bloom control[J]. Chinese Journal of Nature, 2008, 30(5) : 280–286. |
| [29] | May L, Spears B M, Dudley B J, et al. The importance of nitrogen limitation in the restoration of Llangorse Lake, Wales, UK[J]. Journal of Environmental Monitoring, 2010, 12(1) : 338–346. DOI: 10.1039/B912827A |
| [30] | Søndergaard M, Jensen J P, Jeppesen E. Role of sediment and internal loading of phosphorus in shallow lakes[J]. Hydrobiologia, 2003, 506(1-3) : 135–145. |
| [31] | Wang Y C, Wang Z C, Wu W J, et al. Seasonal regime shift of an alternative-state Lake Xingyun, China[J]. Fresenius Environmental Bulletin, 2010, 19(8) : 1474–1485. |
| [32] | 王芸. 洱海夏秋季蓝藻种群动态变化及水华成因分析[J]. 大理学院学报, 2008, 7(12) : 39–42. Wang Y. Study on the group development of cyanophyta and the cause of alga bloom in Erhai Lake in summer and autumn[J]. Journal of Dali University, 2008, 7(12) : 39–42. |
| [33] | Burge K, Breen P F. Detention time design criteria to reduce the risk of excessive algal growth in constructed water bodies[A]. In:Proceedings of the 7th International Conference on Urban Drainage Modelling and the 4th International Conference on Water Sensitive Urban Design[C]. Melbourne:Monash University, 2006. 5-15. |