 2016, Vol. 37
2016, Vol. 37



2. 香港城市大学深圳研究院, 深圳 518057
2. City University of Hong Kong Shenzhen Research Institute, Shenzhen 518057, China
纳米银(AgNPs)是目前应用最广泛、 发展最快的一类纳米材料,由于独特的抗菌特性广泛用于传统行业和一些新兴领域,如涂料、 食品、 纺织、 化妆品、 水质净化、 医疗等[1, 2, 3]. 截止2013年10月,纳米技术消费品目录包含1 628种产品,比2010年增加了24%,其中纳米银相关产品占到383种(www.nanotechproject.org). 纳米银大规模商业化应用在创造巨大经济效益的同时,也存在通过各种途径进入环境的风险,其生物毒性和环境效应一直是人们关注的焦点. Benn等[4]发现市面上6种含有纳米银的袜子洗涤后释放到水中银的浓度高达1 300 μg ·L-1. 一旦纳米银释放进入水环境中,零价态的纳米银可能氧化释放毒性更强的银离子或仍以纳米银的形式存在,造成水体和沉积物持久性污染[5]. 据估计英国每年有8.8 t的纳米银从相关产品中释放进入废水系统,下水道污泥成为一个最主要归宿,当这些污泥用于农业生产时又成为新的污染源[6]. Gottschalk等[7]模型预测结果显示欧洲污水处理厂污泥和河流沉积物中的纳米银含量分别是1 581 ng ·kg-1和952 ng ·kg-1,美国分别是662 ng ·kg-1和195 ng ·kg-1. Chinnapongse等[8]发现一些纳米银能长时间保持稳定最终进入河口淡水或海洋系统,而它们对浮游植物群落的生态毒性并不十分清楚.
纳米银的毒性由多种因素共同决定的,主要包括粒径大小、 包裹物类型、 杂质多少、 表面电荷、 形态结构和银离子溶出速率等[9, 10, 11]. 聚乙烯吡咯烷酮(polyvinyl pyrrolidone)和油胺(oil amine)是稳定性极好的表面活性剂,广泛应用于金、 银等金属纳米材料的制备,纳米银常用的稳定剂有柠檬酸钠(27%)、 聚乙烯吡咯烷酮(PVP,18%)、 胺类(8%)等[12]. Pang等[13]发现纳米银的毒性高度依赖纳米银的表面电荷,其中PVP包裹纳米银的毒性小于柠檬酸钠包裹的纳米银,但大于聚乙二醇包裹的纳米银. 在自然含氧水中,PVP和单宁酸包裹的纳米银比柠檬酸包裹的纳米银更容易释放银离子,而且大粒径的纳米银比小粒径的纳米银更耐溶解并持续释放银离子[5, 14]. Xiu等[15]研究聚乙二醇和PVP包裹的纳米银对大肠杆菌的毒性,发现包裹物主要通过影响银离子释放而间接影响抑菌效果. 大量研究表明纳米银具有潜在的生物毒性作用,并且粒径越小毒性越大[16, 17]. Ivask等[18]发现20-80 nm的纳米银对水生生物的毒性依赖于溶解银离子含量,粒径越大毒性越小;而粒径小于等于10 nm的纳米银因与细胞接触机会更多,具有更高的生物利用度和更强的毒性. Matorin等[19]发现纳米银在低浓度下显著抑制莱茵衣藻光合系统Ⅱ (PSⅡ)电子传递的活性和增加非还原醌的含量,进而抑制光合作用. 虽然纳米银在细胞毒性方面已经有一些研究,但深入比较研究纳米银自身的理化参数以及水环境行为对其毒性影响的较少.
海洋微藻是海洋生态系统的主要初级生产者,具有易培养、 繁殖快、 对环境毒物敏感等优点,常作为实验材料用于化学品毒性测试. 由于适应低光照度、 低温度和广盐性环境,中肋骨条藻广泛分布在全球近海和河口水域[20]. 中肋骨条藻作为浮游植物群落的优势藻种,在适宜条件下极易形成赤潮[21, 22],对研究近海海洋生态环境具有重要意义. 本文在研究纳米银理化参数及其水环境行为的基础上,以河口近岸海域广分布的优势藻中肋骨条藻为受试生物,在细胞和基因两个层次上探讨比较不同包裹物和粒径的纳米银对藻细胞的急性毒性效应,对评价纳米银的生态风险、 监测海洋环境以及赤潮生物种群的变化具有重要意义.
1 材料与方法 1.1 实验仪器与材料仪器:恒温光照培养箱(MGC-800BP-2,上海一恒),电子分析天平(BSA124S,德国Sartorius),超声波清洗仪(SB-520DTDN,宁波新芝),YSI多参数水质分析仪(YSI30,美国维赛),立式压力蒸汽灭菌器(LDZX-50KBS,上海申安),透射电子显微镜(JEM-2100,日本JEOL),动态光散射仪(DLS,Nano-ZS90,英国Malvern),微量紫外分光光度计(SMA4000,美国Merioton),实时定量PCR仪(ABI 7500,美国Applied Biosystems)及其他实验器材.
试剂:10 nm油胺包裹的纳米银粉(型号CST-NP-S10,10 nm-OA)、 10 nm聚乙烯吡咯烷酮包裹的纳米银粉(型号CST-NP-S10,10 nm-PVP)和20 nm聚乙烯吡咯烷酮包裹的纳米银分散在异丙醇制备成10 g ·L-1的纳米银悬浮液(型号CST-NP-S20,20 nm-PVP),3种纳米银均购自苏州冷石纳米材料科技有限公司,纯度为99.5%;其他化学药品和试剂(分析纯)均购自于上海国药集团化学试剂有限公司. 实验的玻璃和塑料器皿使用前用10% HNO3浸泡24 h.
1.2 藻种培养实验藻种为中肋骨条藻(S. costatum,编号CCMA-227),由厦门大学近海海洋国家重点实验室海洋藻类保种中心提供. 藻株于2010年5月采集分离自东海长江口海域,藻种经活化后用于后续实验. 人工海水参照文献[23]的配方配制,充分曝气后,调整pH至7.8-8.4,盐度为28. 经过0.45 μm醋酸纤维酯滤膜过滤后,高压蒸汽灭菌(121℃,20 min),(f/2+Si)海水培养基[24]按照1 ∶1 000的比例将营养液加入到灭菌海水中. 中肋骨条藻按照1 ∶9的比例接种到新鲜培养基中,置于恒温光照培养箱中培养,光强6 000 lx,光暗比14 h ∶10 h,温度(20±1)℃,每日定时摇瓶3次. 暴露实验之前用细胞计数板测定藻细胞密度,并用加富的灭菌海水培养基调整至0.8×109 个 ·L-1.
1.3 纳米银的表征准确称取一定质量的纳米银,将其分散在Milli-Q水和f/2培养基中,40 kHz下超声分散30 min,配制成浓度为5 g ·L-1的悬浮液再稀释到10 mg ·L-1. 取少量分散好的纳米银悬浮液滴在300目的铜网上,37℃恒温干燥2 h后,用透射电子显微镜对纳米银的粒径大小进行表征. 取少量纳米银粉用扫描电子显微镜观察形貌特征,并用动态光散射仪(DLS)测定纳米银的水动力粒径和电动电势.
1.4 藻形态观察纳米银质量浓度设置为0、 5、 50、 500 μg ·L-1一共4组梯度,每组设置3个平行,无菌Milli-Q水作为空白对照. 用移液枪准确移取4 mL对数生长期的藻液到12孔板中,纳米银暴露24 h后,选择浓度为500 μg ·L-1的藻细胞,在光学显微镜下观察藻细胞的形态变化.
1.5 藻细胞存活率取200 μL暴露藻液于96孔酶标板中,加20 μL浓度为5 mg ·mL-1的噻唑蓝(MTT)溶液,室温黑暗孵育4 h后离心5 min,弃上清液,加200 μL二甲亚砜(DMSO)溶解. 用酶标仪在570 nm和630 nm处检测各组的吸光度,对照组吸光度设置为100%,计算藻细胞的存活率.
1.6 ROS含量检测藻细胞内的ROS含量用活性氧检测试剂盒(碧云天生物技术研究所)2,7-二氯荧光素酶(DCFH-DA)法进行测定. 每组分别取200 μL暴露藻液于96孔黑色酶标板中,离心5 min后,弃上清液,加入40 μL的10 μmol ·L-1 DCFH-DA,在37℃孵育30 min. 细胞外的DCFH-DA用f/2培养基洗涤两次,再加入200 μL的f/2培养基,阳性对照组加入等量的氧化剂(Rosup,1 mg ·L-1). 15 min后用酶标仪在488 nm激发波长和525 nm发散波长下检测各组的荧光强度,对照组荧光强度设置为100%,计算ROS含量.
1.7 叶绿素a含量测定釆用90%丙酮萃取叶绿素a,准确移取2 mL藻液于离心管中,4℃、 8 000 r ·min-1离心5 min,弃上清液. 然后每管加入1 mL 90%丙酮,置于4℃冰箱避光萃取24 h后,4℃、 8 000 r ·min-1离心5 min,取上清液,用分光光度计在664 nm和630 nm处测定吸光度值,按照Jeffrey等[25]的公式计算:
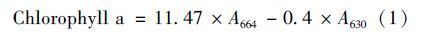
低速离心收集约107个中肋骨条藻细胞的样品,液氮速冻后-80℃冻存,用液氮研磨破碎藻细胞;RNA分离与纯化使用UNlQ-10柱式Trizol总RNA抽提试剂盒(Sangon),用PrimeScriptTM RT reagents with gDNA Eraser试剂盒(TaKaRa)将RNA反转录为cDNA,按照SYBR Premix Ex TaqTM试剂盒(TaKaRa)进行实时荧光定量PCR. 反应程序为:95℃预变性30 s,随后95℃变性5 s和60℃延伸34 s,共40个循环周期,然后用溶解曲线检验产物的专一性. 本研究所用引物由上海生工生物工程有限公司合成,引物序列见表 1. 以 18s 为内参基因,采用2-ΔΔCt方法[26]计算目的基因的相对表达量,即实验组目的基因的表达相对于对照组的表达量的变化倍数.
| 表 1 中肋骨条藻相关基因的引物序列 Table 1 Primer sequences used in gene expression studies of S. costatum |
纳米银的粒径参照比例尺用ImageJ2x软件测量,Excel统计后用Origin 8.0软件制图. 每组实验浓度设置3个平行(n=3),所有实验数据均采用平均值±标准偏差表示. 实验数据用GraphPad Prism 5.0软件进行单因素方差分析及显著性Dunnett-t检验,结果的显著性差异分别以(* P<0.05、 ** P<0.01和*** P<0.001)来表示.
2 结果与分析 2.1 3种纳米银的透射电镜和扫描电镜表征分析图 1分别代表 20 nm-PVP、 10 nm-PVP以及10 nm-OA这3种纳米银分散在Milli-Q水中的透射电镜照片. 图 1(a)显示20 nm-PVP纳米银颗粒由于团聚有重叠且略不规则球形,粒径为20 nm左右;图 1(b)显示10 nm-PVP大部分纳米银颗粒呈椭球形且分散均匀,粒径为10 nm左右;图 1(c)显示10 nm-OA的纳米银呈球形且团聚明显,粒径为10 nm左右. 透射电镜结果显示3种纳米银在Milli-Q水中分散比较均匀,纳米银形态大致呈球形,粒径大小符合正态分布,且粒径越小越容易发生团聚.
 | 图 1 3种纳米银的透射电镜表征 Fig. 1 Representative transmission electron microscopy (TEM) images of three kinds of AgNPs in Milli-Q water |
图 2分别为20 nm-PVP、 10 nm-PVP以及10 nm-OA这3种纳米银在干粉状态下的扫描电镜图. 图 2(a)显示20 nm-PVP的纳米银呈灰白色椭球形,大粒径占绝大多数;图 2(b)显示10 nm-PVP的纳米银呈灰白色球形,粒径较小且大小基本一致;图 2(c)显示10 nm-OA纳米银颗粒团聚在一起呈块状并被包裹物完全覆盖. 从纳米银颗粒的形貌特征分析,10 nm-PVP和20 nm-PVP拥有相同的包裹物,类型明显不同于10 nm-OA的包裹物. 扫描电镜结果说明PVP包裹的纳米银比油胺包裹的纳米银分散性更好.
 | 图 2 3种纳米银的扫描电镜表征 Fig. 2 Representative scanning electron microscopy (SEM) images of three kinds of AgNPs powder |
选取3种纳米银在Milli-Q水分散介质中有代表性的透射电镜图,每种纳米银的粒径分布分别统计140个纳米银颗粒的直径. 结果如下:10 nm-OA粒径为(8.27±1.80) nm;20 nm-PVP粒径为(11.20±3.44) nm;10 nm-PVP粒径为(18.20±3.98) nm. 3种纳米银的粒径大小符合正态分布(图 3),10 nm的两种纳米银中值粒径略有差异,但与20 nm-PVP中值粒径相比差异显著.
 | 图 3 3种纳米银在f/2培养基中的粒径分布 Fig. 3 Size distribution of three kinds of AgNPs in f/2 medium |
表 2列出了3种纳米银在Milli-Q水和f/2培养基中的水动力粒径和电动电势变化. 纳米银在f/2培养基中水动力粒径明显大于Milli-Q水中,10 nm-PVP和10 nm-OA两种纳米银的水动力粒径较大且比较接近,而20 nm-PVP在f/2培养基中的水动力粒径变化不大,说明纳米银粒径越小越容易发生团聚. 其中10 nm-PVP的中值粒径最大为142 nm;而20 nm-PVP在两种分散介质中的水动力粒径变化不大,在f/2培养基中的水动力粒径明显小于10 nm的两种纳米银.
| 表 2 3种纳米银在水介质中的水动力粒径及其电动电势 Table 2 The z-average hydrodynamic diameter and zeta potential of the three kinds of AgNPs in water |
3种纳米银在Milli-Q水中的电动电势明显小于在f/2培养基中,其中10 nm-PVP在Milli-Q水中的电动电势最小为(-39.8±7.71)mV,而在f/2培养基中3种纳米银的电动电势趋于一致. 纳米银在两种介质中的电动电势与水动力粒径变化趋势一致,由于电动电势越低粒子越不稳定,说明纳米银在f/2培养基中发生不同程度的团聚. 电镜表征结果显示10 nm的两种纳米银的中值粒径略有差异,但这两种纳米银在f/2培养基中的水动力粒径比较接近,说明包裹物对纳米银在水介质中粒径大小有不同影响,进而间接影响纳米银的毒性.
2.3 纳米银粒径和包裹物对藻细胞存活率的影响中肋骨条藻暴露于3种纳米银24 h后,藻细胞的存活率随着纳米银浓度的升高而减少(图 4). 在纳米银浓度≥5 μg ·L-1时,10 nm-OA处理组藻细胞存活率显著减少(P<0.001);在纳米银浓度≥500 μg ·L-1时,10 nm-PVP和20 nm-PVP处理组藻细胞存活率都显著减少(P<0.001). 相同粒径不同包裹物的两种纳米银在对藻细胞毒性方面,10 nm-OA的毒性大于10 nm-PVP. 说明PVP比油胺更能降低纳米银的毒性. 而相同包裹物的两种纳米银对藻细胞毒性随纳米银浓度不同而差异很大. 在纳米银浓度小于500 μg ·L-1时,20 nm-PVP的毒性大于10 nm-PVP;在纳米银浓度≥500 μg ·L-1时,20 nm-PVP的毒性小于10 nm-PVP,说明纳米银的毒性大小取决于纳米银的水动力粒径和暴露浓度.
 | 图 4 不同包裹物和粒径的纳米银对中肋骨条藻细胞存活率的影响 Fig. 4 Cell viability of S. costatum upon exposure to AgNPs with different surface coatings and particle sizes for 24 h |
3种纳米银对藻细胞存活率结果表明,纳米银在500 μg ·L-1时能显著降低藻细胞的存活率. 图 5显示了对照组和500 μg ·L-1不同纳米银处理组藻细胞的形态学变化,红色箭头标注为死亡藻细胞的硅质外壳,黄色箭头标注为死亡的藻细胞碎片. 结果显示10 nm-PVP和10 nm-OA纳米银毒性大于20 nm-PVP纳米银,死亡的藻细胞数量也更多,并且出现大量死亡的藻细胞碎片. 在对藻细胞形态损伤方面,粒径大小是纳米银毒性的主要因素,而且粒径越小毒性越大.
 | 图 5 中肋骨条藻在500 μg ·L-1纳米银暴露下形态学变化 Fig. 5 Cell morphological changes of S. costatum after exposure to 500 μg ·L-1 AgNPs for 24 h |
中肋骨条藻暴露于PVP包裹的两种纳米银24 h后,藻细胞内活性氧的含量随着纳米银的浓度升高而减少(图 6). 在浓度≥500 μg ·L-1时,10 nm-PVP处理的藻细胞内活性氧含量显著减少(P<0.01);在浓度≥50 μg ·L-1时,20 nm-PVP处理的藻细胞内活性氧含量显著减少(P<0.05). 说明PVP包裹的纳米银能够减少对藻细胞的氧化损伤,而且纳米银粒径越小毒性减小得越多. 此外,中肋骨条藻暴露于10 nm油胺包裹的纳米银24 h后,藻细胞内活性氧的含量随着纳米银的浓度升高而增加,在浓度≥500 μg ·L-1时,藻细胞内活性氧含量显著升高(P<0.001). 油胺和PVP包裹的纳米银对藻细胞氧化应激和氧化损伤效应结果表明,10 nm油胺包裹的纳米银通过诱导藻细胞产生过多的活性氧而具有更强的细胞毒性.
 | 图 6 不同包裹物和粒径的纳米银对中肋骨条藻活性氧的影响 Fig. 6 Induction of ROS in S. costatum after exposed to AgNPs with different surface coatings and particle sizes for 24 h |
中肋骨条藻暴露于10 nm-PVP和10 nm-OA纳米银24 h后,10 nm-OA处理组藻细胞叶绿素a含量随着纳米银的浓度升高而增加,在浓度≥50 μg ·L-1时,叶绿素a含量显著升高(P<0.05). 中肋骨条藻暴露于10 nm-PVP和20 nm-PVP纳米银 24 h后,藻细胞叶绿素a含量随着纳米银浓度的升高而减少(图 7). 在浓度≥50 μg ·L-1时,10 nm-PVP处理组藻细胞叶绿素a含量显著减少(P<0.01);在浓度≥500 μg ·L-1时,20 nm-PVP处理组藻细胞叶绿素a含量显著减少(P<0.01). 而10 nm-OA和10 nm-PVP对藻细胞叶绿素a含量影响差异显著,10 nm-OA处理组藻细胞叶绿素a含量增加,10 nm-PVP则降低了藻细胞叶绿素a含量. 结果表明相同包裹物条件下,纳米银粒径越小藻细胞叶绿素a含量减少得越明显.
 | 图 7 不同包裹物和粒径的纳米银对中肋骨条藻叶绿素a含量的影响 Fig. 7 Change of chlorophyll a content in S. costatum upon exposure to AgNPs with different surface coatings and particle sizes for 24 h |
中肋骨条藻暴露于3种纳米银24 h后,结果表明10 nm-PVP和10nm-OA处理组藻细胞光合作用基因的表达量变化显著,而20 nm-PVP处理组没有显著性变化(图 8). 3HfcpA 基因随着纳米银浓度升高有上调的趋势,10 nm-OA处理组在浓度≥500 μg ·L-1时显著上调(P<0.01),而10 nm-PVP处理组在浓度≥50 μg ·L-1时显著上调(P<0.05). 3HfcpB 与 3HfcpA 基因是同源基因,其变化与 3HfcpA 基因相反,随着10 nm-PVP和10 nm-OA浓度升高有下调的趋势. 中肋骨条藻PSⅡ中心反应蛋白D 1 基因的表达明显受到纳米银的抑制,10 nm-PVP和10 nm-OA处理组藻细胞D 1 基因随纳米银浓度的升高有下调的趋势,其中10 nm-PVP在浓度≥50 μg ·L-1时显著下调(P<0.05). 纳米银粒径越小对光合作用相关基因表达的影响越显著,而相同粒径条件下,PVP比油胺包裹的纳米银更显著诱导藻细胞光合作用基因变化,说明小粒径PVP包裹的纳米银对光合作用基因的影响最大.
 | 图 8 不同包裹物和粒径的纳米银对中肋骨条藻光合作用相关基因表达水平的影响 Fig. 8 Expression changes of photosynthesis relevant gene in S. costatum following exposure to AgNPs with different surface coatings and particle sizes for 24 h |
影响纳米银在水环境中稳定性的因素都会影响到纳米银对藻类的细胞毒性,这些因素不仅包括材料本身特性(形状大小、 包裹物类型、 贮存状态),还与分散剂和环境因素(pH、 离子强度、 离子成分、 溶解性有机物等)有关[9, 29]. 3种纳米银的电镜表征结果表明,纳米银颗粒呈椭球形且大小均匀,因而具有极大的比表面积. 另外,DLS数据表明纳米银在Milli-Q水和f/2培养基两种介质中均有不同程度的团聚,水动力粒径和电动电势差别很大. 许多纳米材料由于大的比表面积和强的吸附能力,极易发生团聚或吸附在藻细胞表面,形成物理遮蔽或细胞接触而产生毒性,如纳米TiO2[30]. 纳米银的水动力粒径随着水环境中离子浓度升高而粒径增大,电动电势也表现出相同的趋势(表 2),从侧面证实纳米银团聚后稳定性降低. 由于20 nm-PVP分散在异丙醇溶剂中储存,因而具有良好的分散效果,在水环境介质中水动力粒径变化不大,在一定程度上增强纳米银的毒性. 而10 nm-PVP和20 nm-PVP两种纳米银在f/2培养基中的水动力粒径大小和电动电势的明显差异,说明小粒径的纳米银颗粒之间的相互作用力更强,在海水环境中更容易发生团聚,这在一方面解释了纳米银的团聚行为对藻细胞毒性的影响. 此外,纳米银的浓度和粒径大小对藻细胞毒性具有条件限制作用,当纳米银浓度小于500 μg ·L-1时,大粒径的纳米银毒性更大;在浓度≥500 μg ·L-1时,小粒径的纳米银毒性更大. 本研究主要在氧化损伤和光合作用两个方面比较了3种纳米银对藻细胞的毒性差异,纳米银对藻细胞的可能毒性机制还有溶解释放银离子[31]和生物富集[32]等途径,这有待进一步研究.
3.2 纳米银对藻细胞的氧化损伤效应通过比较3种纳米银对藻细胞存活率和形态的影响,发现中肋骨条藻细胞存活率与细胞内活性氧的积累呈负相关,说明氧化损伤效应是纳米银对藻细胞毒性的主要途径之一. 大量研究结果也证实纳米银是通过诱导产生过多的活性氧发挥细胞毒性并抑制藻细胞的生长[33, 34]. 3种纳米银的毒性大小依次为10 nm-OA>10 nm-PVP>20 nm-PVP,说明纳米银的毒性主要取决于粒径大小. 在对藻细胞氧化损伤方面,发现油胺包裹的纳米银显著诱导藻细胞产生过多的活性氧,而PVP包裹的纳米银处理组藻细胞内活性氧水平有显著性减少,说明PVP能够降低纳米银的生物毒性. 关于PVP包裹纳米银的大量毒理研究表明[35, 36],PVP包裹的纳米银较柠檬酸包裹的纳米银化学结构更稳定,而且在环境介质中银的溶解速率更小,因而具有更小的生物毒性,说明PVP包裹的纳米银更安全. 在对藻细胞叶绿素a含量影响方面,10 nm-PVP和10 nm-OA在50 μg ·L-1下引起叶绿素a含量显著性变化,而20 nm-PVP处理组在500 μg ·L-1下出现显著性变化,说明小粒径纳米银对藻细胞叶绿素a含量影响显著,叶绿素可能是纳米银毒性的重要靶点. Li等[30]对纳米TiO2研究已经证实藻细胞叶绿体是活性氧产生和积累的位点,这在另一方面说明纳米银通过影响叶绿素a含量和活性氧含量并影响光合作用过程.
3.3 纳米银对藻细胞光合作用的毒性影响光合作用是藻类最重要的生命活动,目前对藻类在不同纳米银下分子水平的基因表达变化情况所知甚少. 在对中肋骨条藻光合作用相关基因表达的研究发现,3种纳米银处理下藻细胞叶绿素捕光蛋白基因 3HfcpA 、 3HfcpB 和PSⅡ的中心反应蛋白基因D 1 均出现显著性变化,10 nm-PVP和10 nm-OA两种纳米银对光合作用基因的干扰最严重,而20 nm-PVP处理组没有显著性变化. 同源基因 3HfcpA 和 3HfcpB 主要参与光反应中捕光蛋白的合成[27], 3HfcpA 基因的上调表达引起 3HfcpB 基因的下调表达,这可能是藻细胞在受到纳米银胁迫后的一种光保护机制,说明纳米银影响了藻细胞对光的吸收过程. 10 nm-PVP和10nm-OA处理组藻细胞D 1 基因表达显著下调,说明PSⅡ的中心反应蛋白D1合成受阻. Ali等[37]研究表明重金属铬诱导PSⅡ功能活性的变化和活性氧的形成与D1蛋白的含量减少高度相关. 也有研究发现植物细胞在强光胁迫下可能引起D1蛋白的损伤[38],D1蛋白的破坏可能导致PSⅡ反应中心结构和功能的损害,继而引起光合电子传递的受阻,进而抑制光合作用的正常进行[39]. 因此,纳米银对藻细胞的另外一个毒性途径可能是通过调控光合作用关键蛋白质的合成,干扰光吸收和PSⅡ电子传递等过程,进而抑制光合系统活性和藻细胞生长. 本研究表明,纳米银对中肋骨条藻的细胞毒性和基因毒性呈现一致性,并且粒径大小是纳米银毒性的主要因素,而且PVP这种包裹物比油胺更能降低纳米银的毒性. 关于PVP和油胺这两种包裹物与纳米银的结合方式以及如何与藻细胞相互作用的机制方面还需要深入研究.
4 结论(1)纳米银对中肋骨条藻的生长有抑制作用并呈现明显的剂量效应关系,其毒性大小依次为10 nm-OA>10 nm-PVP>20 nm-PVP.
(2)纳米银的粒径越小对中肋骨条藻的细胞毒性越大,此外,水环境中纳米银的粒径和有效浓度是影响其毒性的重要因素.
(3)包裹物通过影响纳米银在水环境中的稳定性间接影响纳米银的毒性,其中聚乙烯吡咯烷酮(PVP)比油胺(OA)更能降低纳米银的生物毒性.
(4)通过比较3种纳米银对中肋骨条藻细胞毒性的影响,发现藻细胞存活率与细胞内活性氧的积累呈负相关关系,而且小粒径纳米银显著诱导D 1 基因下调表达进而抑制光合作用,说明活性氧介导的氧化损伤和对光合作用关键基因调控是纳米银对藻细胞可能的毒性作用机制.
| [1] | Lalley J, Dionysiou D D, Varma R S, et al. Silver-based antibacterial surfaces for drinking water disinfection-an overview[J]. Current Opinion in Chemical Engineering, 2014, 3: 25-29. |
| [2] | Tang B, Wang J F, Xu S P, et al. Application of anisotropic silver nanoparticles: multifunctionalization of wool fabric[J]. Journal of Colloid and Interface Science, 2011, 356 (2): 513-518. |
| [3] | Chaloupka K, Malam Y, Seifalian A M. Nanosilver as a new generation of nanoproduct in biomedical applications[J]. Trends in Biotechnology, 2010, 28 (11): 580-588. |
| [4] | Benn T M, Westerhoff P. Nanoparticle silver released into water from commercially available sock fabrics[J]. Environmental Science & Technology, 2008, 42 (11): 4133-4139. |
| [5] | Dobias J, Bernier-Latmani R. Silver release from silver nanoparticles in natural waters[J]. Environmental Science & Technology, 2013, 47 (9): 4140-4146. |
| [6] | Whiteley C M, Valle M D, Jones K C, et al. Challenges in assessing release, exposure and fate of silver nanoparticles within the UK environment[J]. Environmental Science: Processes & Impacts, 2013, 15 (11): 2050-2058. |
| [7] | Gottschalk F, Sonderer T, Scholz R W, et al. Modeled environmental concentrations of engineered nanomaterials (TiO2, ZnO, Ag, CNT, Fullerenes) for different regions[J]. Environmental Science & Technology, 2009, 43 (24): 9216-9222. |
| [8] | Chinnapongse S L, MacCuspie R I, Hackley V A. Persistence of singly dispersed silver nanoparticles in natural freshwaters, synthetic seawater, and simulated estuarine waters[J]. Science of the Total Environment, 2011, 409 (12): 2443-2450. |
| [9] | Fabrega J, Luoma S N, Tyler C R, et al. Silver nanoparticles: behaviour and effects in the aquatic environment[J]. Environment International, 2011, 37 (2): 517-531. |
| [10] | Park J W, Oh J H, Kim W K, et al. Toxicity of citrate-coated silver nanoparticles differs according to method of suspension preparation[J]. Bulletin of Environmental Contamination and Toxicology, 2014, 93 (1): 53-59. |
| [11] | Burchardt A D, Carvalho R N, Valente A, et al. Effects of silver nanoparticles in diatom Thalassiosira pseudonana and cyanobacterium Synechococcus sp.[J]. Environmental Science & Technology, 2012, 46 (20): 11336-11344. |
| [12] | Tolaymat T M, Badawy A M E, Genaidy A, et al. An evidence-based environmental perspective of manufactured silver nanoparticle in syntheses and applications: a systematic review and critical appraisal of peer-reviewed scientific papers[J]. Science of the Total Environment, 2010, 408 (5): 999-1006. |
| [13] | Pang C F, Brunelli A, Zhu C H, et al. Demonstrating approaches to chemically modify the surface of Ag nanoparticles in order to influence their cytotoxicity and biodistribution after single dose acute intravenous administration[J]. Nanotoxicology, 2015, doi: 10.3109/17435390.2015.1024295. |
| [14] | Levard C, Hotze E M, Lowry G V, et al. Environmental transformations of silver nanoparticles: impact on stability and toxicity[J]. Environmental Science & Technology, 2012, 46 (13): 6900-6914. |
| [15] | Xiu Z M, Zhang Q B, Puppala H L, et al. Negligible particle-specific antibacterial activity of silver nanoparticles[J]. NANO Letters, 2012, 12 (8): 4271-4275. |
| [16] | Carlson C, Hussain S M, Schrand A M, et al. Unique cellular interaction of silver nanoparticles: size-dependent generation of reactive oxygen species[J]. The Journal of Physical Chemistry B, 2008, 112 (43): 13608-13619. |
| [17] | Kim T H, Kim M, Park H S, et al. Size-dependent cellular toxicity of silver nanoparticles[J]. Journal of Biomedical Materials Research Part A, 2012, 100 (4): 1033-1043. |
| [18] | Ivask A, Kurvet I, Kasemets K, et al. Size-dependent toxicity of silver nanoparticles to bacteria, yeast, algae, crustaceans and mammalian cells in vitro[J]. PLoS One, 2014, 9 (7): e102108. |
| [19] | Matorin D N, Todorenko D A, Seifullina N K, et al. Effect of silver nanoparticles on the parameters of chlorophyll fluorescence and P700 reaction in the green alga Chlamydomonas reinhardtii[J]. Microbiology, 2013, 82 (6): 809-814. |
| [20] | Liu L S, Zhou J, Zheng B H, et al. Temporal and spatial distribution of red tide outbreaks in the Yangtze River Estuary and adjacent waters, China[J]. Marine Pollution Bulletin, 2013, 72 (1): 213-221. |
| [21] | Wang Q, Zhu L S, Hu J P. A HAB mathematical model of competition between two algae species in the Yangtze River Estuary and its adjacent waters[A]. In: Proceedings of 2010 International Conference on Multimedia Technology (ICMT)[C]. Ningbo: IEEE, 2010. 1-4. |
| [22] | 杨世民, 王丽莎, 石晓勇. 2009年夏季胶州湾同步调查浮游植物群落结构特征[J]. 海洋与湖沼, 2015, 46 (1): 102-108. |
| [23] | Kester D R, Duedall I W, Connors D N, et al. Preparation of artificial seawater[J]. Limnology and Oceanography, 1967, 12 (1): 176-179. |
| [24] | Guillard R R L, Ryther J H. Studies of marine planktonic diatoms. I. cyclotella nana Hustedt, and Detonula confervacea (cleve) Gran[J]. Canadian Journal of Microbiology, 1962, 8 (2): 229-239. |
| [25] | Jeffrey S T, Humphrey G F. New spectrophotometric equations for determining chlorophylls a, b, c1 and c2 in higher plants, algae and natural phytoplankton[J]. Biochemie und Physiologie der Pflanzen (BPP), 1975, 167: 191-194. |
| [26] | Kim K T, Zaikova T, Hutchison J E, et al. Gold nanoparticles disrupt zebrafish eye development and pigmentation[J]. Toxicological Sciences, 2013, 133 (2): 275-288. |
| [27] | Bopp S K, Lettieri T. Gene regulation in the marine diatom Thalassiosira pseudonana upon exposure to polycyclic aromatic hydrocarbons (PAHs)[J]. Gene, 2007, 396 (2): 293-302. |
| [28] | Kim Tiam S, Feurtet-Mazel A, Delmas F, et al. Development of q-PCR approaches to assess water quality: effects of cadmium on gene expression of the diatom Eolimna minima[J]. Water Research, 2012, 46 (4): 934-942. |
| [29] | 苑志华, 唐婷, 于昌平. 不同封端剂纳米银对氨氧化细菌的毒性研究[A]. 见: 第六届全国环境化学学术大会暨环境科学仪器与分析仪器展览会摘要集[C]. 北京: 中国化学会, 2011. |
| [30] | Li F M, Liang Z, Zheng X, et al. Toxicity of nano-TiO2 on algae and the site of reactive oxygen species production[J]. Aquatic Toxicology, 2015, 158: 1-13. |
| [31] | Navarro E, Wagner B, Odzak N, et al. Effects of differently coated silver nanoparticles on the photosynthesis of Chlamydomonas reinhardtii[J]. Environmental Science & Technology, 2015, 49 (13): 8041-8047. |
| [32] | Leclerc S, Wilkinson K J. Bioaccumulation of nanosilver by Chlamydomonas reinhardtii-nanoparticle or the free ion?[J]. Environmental Science & Technology, 2014, 48 (1): 358-364. |
| [33] | Oukarroum A, Barhoumi L, Pirastru L, et al. Silver nanoparticle toxicity effect on growth and cellular viability of the aquatic plant Lemna gibba[J]. Environmental Toxicology and Chemistry, 2013, 32 (4): 902-907. |
| [34] | Oukarroum A, Bras S, Perreault F, et al. Inhibitory effects of silver nanoparticles in two green algae, Chlorella vulgaris and Dunaliella tertiolecta[J]. Ecotoxicology and Environmental Safety, 2012, 78: 80-85. |
| [35] | Lu W T, Senapati D, Wang S G, et al. Effect of surface coating on the toxicity of silver nanomaterials on human skin keratinocytes[J]. Chemical Physics Letters, 2010, 487 (1-3): 92-96. |
| [36] | Angel B M, Batley G E, Jarolimek C V, et al. The impact of size on the fate and toxicity of nanoparticulate silver in aquatic systems[J]. Chemosphere, 2013, 93 (2): 359-365. |
| [37] | Ali N A, Dewez D, Didur O, et al. Inhibition of photosystem Ⅱ photochemistry by Cr is caused by the alteration of both D1 protein and oxygen evolving complex[J]. Photosynthesis Research, 2006, 89 (2-3): 81-87. |
| [38] | Domingues N, Matos A R, da Silva J M, et al. Response of the diatom Phaeodactylum tricornutum to photooxidative stress resulting from high light exposure[J]. PLoS One, 2012, 7 (6): e38162. |
| [39] | 郭延平, 刘辉, 胡美君, 等. 植物PSⅡ反应中心D1蛋白的结构与功能[A]. 见: 中国园艺学会第七届青年学术讨论会论文集[C]. 北京: 中国园艺学会, 2006. |