 2016, Vol. 37
2016, Vol. 37



2. 国家海洋局第一海洋研究所, 国家海洋局海洋生物活性物质重点实验室, 青岛 266061
2. Key Laboratory of Marine Bioactive Substances, First Institute of Oceanography, State Oceanic Administration, Qingdao 266061, China
异养硝化-好氧反硝化 (heterotrophic nitrification-aerobic denitrification,HN-AD)菌是一类能够在有机物存在的条件下将氨氮氧化、 在溶解氧存在条件下将亚硝态氮、 硝态氮代谢为氮气的微生物. 早在1983年,Robertson等[1]就从脱硫脱氮污水处理系统中分离出1株具有HN-AD能力的副球菌Paracoccus pantotrophus ATCC 35512(曾被命名为Thiosphaera pantotropha). 此后,具有此特性的菌株被从不同环境中不断分离出来[2, 3, 4, 5, 6, 7, 8, 9]. 由于异养硝化-好氧反硝化菌具有生长快速、 耐有机负荷、 耐溶解氧、 脱氮效率高等特点,在污水处理中具有潜在的优势,成为当今的研究热点.
目前,许多国家和地区都在加快对海水的直接利用以缓解所面临的缺水问题. 海水直接利用产生的废水盐度普遍较高,而高盐环境会导致微生物的生长代谢受到抑制,脱氢酶活性降低,较高的渗透压还会引起微生物细胞的质壁分离,导致细胞破裂死亡[10]. 国内外对HN-AD菌的研究多集中在生活污水的处理中,对同时具有耐盐和HN-AD功能的菌株的研究鲜见报道.
本研究从胶州湾海底沉积物中分离筛选得到1株海洋HN-AD菌y5,对其进行了鉴定. 探讨了高盐环境下不同有机碳源、 初始pH值以及不同C/N值对该菌株生长及其脱氮性能的影响. 通过研究其在高盐单一或混合氮源条件下的脱氮特性,以期丰富生物脱氮理论并且为开发高盐废水的生物处理工艺提供一定的理论和技术支持.
1 材料与方法 1.1 菌株来源和培养基本实验所用菌株是从胶州湾海底沉积物中分离筛选得到.
菌株分离和生长培养基:KH2PO4 0.2 g,K2SO4 0.1 g,NH4Cl 0.5 g,KNO2 1 g,柠檬酸三钠 1.5 g,微量元素溶液Ⅰ、 Ⅱ各1.25 mL,海水1 000 mL,pH 7.0-8.0. 固体培养基加入2%琼脂粉.
观察菌落的形态学特征和活化菌株时采用海水2216E培养基:蛋白胨 5 g,酵母粉 1 g,海水1 000 mL,固体培养基加入2%琼脂粉.
微量元素溶液Ⅰ:EDTA 5 g ·L-1,FeSO4 ·7H2O 5 g ·L-1.
微量元素溶液Ⅱ:EDTA 15 g ·L-1,H3BO4 0.014 g ·L-1,MnCl2 ·4H2O 0.99 g ·L-1,CuSO4 ·5H2O 0.25 g ·L-1,ZnSO4 ·7H2O 0.43 g ·L-1,NiCl ·6H2O 0.19 g ·L-1,NaMoO4 ·2H2O 0.22 g ·L-1,CoCl2 ·6H2O 0.24 g ·L-1,NaSeO4 ·10H2O 0.21 g ·L-1.
1.2 菌株的鉴定 1.2.1 细菌形态观察和生理生化实验将菌株接种至2216E海水固体培养基上,观察菌株在固体培养基上的菌落特征. 通过拍摄扫描电镜照片来观察菌株的个体形态. 利用细菌生化鉴别试剂盒(BIO-KNOT Rap ·E-15肠杆菌科鉴定) 对菌株进行生化鉴定.
1.2.2 16S rRNA测序及系统发育分析接种一环菌株至液体海水2216E培养基,以温度28℃、 转速160 r ·min-1培养12 h后取200 μL菌液于EP管中,先经沸水浴13 min再经冰浴7 min后制得DNA模板. 通过正向引物27F:5′-AGAGTTTGATCCTGGTCAG-3′,反向引物1492R:5′-GGTTACCTTGTTACGACTT-3′进行PCR扩增.
PCR反应体系(50 μL):正向引物27F(10 μmol ·L-1) 1 μL; 反向引物1492R(10 μmol ·L-1)1 μL; 2×Tap PCR MasterMix 25 μL; 细菌DNA模板 2 μL; ddH2O 21 μL.
PCR反应程序:95℃预变性5 min,94℃变性 40 s,52℃ 复性40 s,72℃ 延伸90 s,30 个循环,72℃延伸 7 min.
系统发育分析:PCR 产物由上海金斯瑞生物有限公司测序,测序结果与 GenBank中已提交的模式菌株的16S rRNA 基因序列通过Blast软件进行同源性比较,通过BIOEDIT软件中的程序进行多重序列比对分析,利用MEGA6.0软件,以Neighbor-joining法构建系统发育树.
1.3 菌株脱氮特性影响因素研究如表 1 所示,选择的因素为碳源、 pH、 C/N三项,以分离培养基为基础,调节培养基的初始pH值为7.0(2号除外),C/N为10(3号除外),按2%的接种量即吸取对数生长期的菌液(D600约为0.80)2 mL接种至100 mL灭菌后的培养基中. 置于气浴恒温振荡器(转速160 r ·min-1)中,28℃ 培养24 h后检测培养基中菌体的生长量(D600),氨氮、 亚硝态氮和硝态氮的浓度.
|
|
表 1 菌株y5生长条件实验设置 Table 1 Experimental setting of strain y5 growth conditions |
以分离培养基为基础,柠檬酸三钠为碳源,pH=7.0,C/N为17配制菌株y5的高盐脱氮测试液. 分别以氯化铵、 亚硝酸钠、 硝酸钾为唯一氮源,研究菌株y5的异养硝化作用和好氧反硝化作用. 分别配制含氯化铵+亚硝酸钠、 氯化铵+硝酸钾、 氯化铵+亚硝酸钠+硝酸钾的降解测试液,研究菌株y5在混合氮源系统中的异养硝化-好氧反硝化作用.
将活化后处于对数增长期的菌液(D600约为0.80)按2%的接种量接入盛有高温灭菌后的300 mL测试液的500 mL三角瓶中,实验设置2个重复,置于恒温气浴振荡器(转速160 r ·min-1)中,28℃ 培养,定时取样,测定测试液中的菌体的生长量(D600),氨氮、 亚硝态氮和硝态氮浓度.
1.5 分析方法对氨氮、 亚硝态氮、 硝态氮浓度以及菌体生长量(D600)的测定均采用UV 5200紫外/可见分光光度计进行. 氨氮:钠氏试剂分光光度法; 亚硝态氮:N-(1-萘基)乙二胺分光光度法; 硝态氮:麝香草酚分光光度法; 菌体生长量:测定菌液在600 nm波长时的吸光度值(D600),pH和ORP使用WTW pH/Oxi340i/SET测定. 脱氮率计算方法如下:
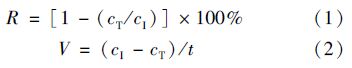
菌株y5在固体培养基上培养24 h后,菌落大小约为1.0-1.5 mm,呈圆形,乳白色,表面光滑,湿润,隆起,不透明,黏稠,边缘整齐[如图 1(a)]. 通过扫描电子显微镜观察菌体如图 1(b)所示,菌株为短杆菌,大小为0.5 μm×0.2 μm,无芽孢和鞭毛. 对菌株y5生理生化鉴定的结果表明,该菌为不动杆菌,氧化酶、 水解酶反应阳性,甲基红实验、 吲哚实验阴性. 菌株y5的生理生化鉴定结果见表 2.

|
图 1 菌株y5的形态特征 Fig. 1 Electronic microscope photograph of strain y5 |
|
|
表 2 菌株y5的生理生化特征 Table 2 Physiological and biochemical characteristics of strain y5 |
对菌株y5测序获得长度为1 363 bp的部分16S rRNA基因序列,在GenBank中 的 登 录 号:KP215627. 将所得的基因序列提交至GenBank中通过Blast检索,应用MEGA 6.0软件,以Neighbor-joining 法绘制系统发育树,如图 2所示. 菌株y5与菌株Klebsiella pneumoniae AUH-BG208的同源性为100%,与多株Klebsiella pneumoniae 同源性达99%,结合菌株的形态特征和生理生化特征,可初步确定菌株y5属克雷伯氏菌属(Klebsiella sp.).

|
图 2 基于16S rRNA基因序列同源性构建的菌株y5的系统发育树 Fig. 2 Phylogenetic tree of strain y5 based on the complete sequence of 16S rRNA gene |
如图 3所示,菌株y5在以乙酸钠、 丁二酸钠、 柠檬酸三钠、 葡萄糖和蔗糖为唯一碳源时均能生长. 以蔗糖为唯一碳源时,菌株y5的D600值最高,生长最好,其次为葡萄糖、 柠檬酸三钠. 碳源为柠檬酸三钠时,菌株y5对氨氮的去除效率最高为71.02%,对硝态氮的去除率最高为90.72%. 蔗糖和葡萄糖为碳源时,对硝态氮的去除率也比较高,但是系统内有大量的亚硝态氮的积累,氨氮的去除率也较低. 所以菌株y5的最佳碳源是柠檬酸三钠.

|
图 3 碳源对菌株y5生长和脱氮反应的影响 Fig. 3 Effects of carbon resource on the growth and nitrogen removal reaction of strain y5 |
初始pH为培养基灭菌后的pH,分别为4.9、 5.4、 5.9、 6.4、 7.0、 7.4、 8.0、 8.4、 8.9、 9.5. 如图 4所示,反应24 h以后,初始pH为6.0~8.0时硝态氮的去除率都在99%以上,菌体生长较好. 当pH为7.0时,硝态氮的脱除率最大达到100%,D600值最大为1.09,亚硝态氮的累积量也较低. pH为7.0~7.5时,氨氮的去除率都在60%以上. 当pH为7.0时,氨氮的去除率最大为65.24%. 所以菌株y5的最适pH为7.0.

|
图 4 不同pH对菌株y5生长和脱氮反应的影响 Fig. 4 Effects of pH on the growth and nitrogen removal reaction of strain y5 |
如图 5所示,随着C/N的升高,菌株y5的生长量、 对氨氮和硝态氮的脱除率也随之升高,当C/N=17时,菌株的生长量达到最大值为1.09,氨氮和硝态氮的脱除率也达到最大值分别为99.86%和85.88%,之后均有所下降. 当C/N比小于5时,菌株y5的生长量较低,对氨氮和硝态氮的脱除效果也较差. 在C/N为9-21的范围内,菌株的反硝化能力均在97%以上,当C/N为9~17时,氨氮的降解率均在79%以上. 菌株y5的最佳C/N为17.

|
图 5 不同C/N对菌株y5生长和脱氮反应的影响 Fig. 5 Effects of C/N ratio on the growth and nitrogen removal reaction of strain y5 |
如图 6所示,以NH4Cl为唯一氮源时,0-9 h为菌株的生长适应期,菌体从9 h开始进入对数增长期,D600最大为1.32. NH4+-N的浓度由初始的100.75 mg ·L-1,经36 h反应后,降至22.47 mg ·L-1,去除率为77.07%. NH4+-N从9 h开始快速去除,平均去除速率为2.59 mg ·(L ·h)-1,这说明NH4+-N的去除主要发生在细菌的对数增长期. 反应过程中,几乎检测不到硝酸盐和亚硝酸盐的积累.

|
图 6 菌株y5的异养硝化过程 Fig. 6 Process of heterotrophic nitrification of strain y5 |
如图 7所示,以NaNO2为唯一氮源时,0-12 h菌株y5处于适应期,12 h以后进入对数增长期,D600最大为0.70. NO2--N的浓度由初始的189.15 mg ·L-1,经36 h反应后,降至67.99 mg ·L-1,去除率为64.14%. NO2--N在15 h以后进入快速降解阶段,平均降解速率为5.21 mg ·(L ·h)-1. 说明菌株y5的亚硝酸盐好氧反硝化作用主要发生在对数增长期. 亚硝酸盐的降解过程中,没有明显的氨氮和硝酸盐氮的累积.

|
图 7 菌株y5的亚硝酸盐好氧反硝化过程 Fig. 7 Process of nitrate denitrification of strain y5 |
如图 8所示,以KNO3为唯一氮源时,0-12 h为菌株y5的生长适应期,12 h以后进入对数增长期,D600最大为1.03,在21-27 h内菌体的生长处在停滞状态,这是因为亚硝态氮的大量累积对细菌的生长产生了抑制. NO3--N的浓度由初始的86.71 mg ·L-1,经36 h反应后,降至0,去除率为100%. 12-24 h为NO3--N的快速降解阶段,平均降解效率为6.37 mg ·(L ·h)-1,菌株y5的好氧反硝化作用主要发生在对数增长期. NO3--N的还原过程中,产生的NO2--N在24 h时累积量最大为33.13 mg ·L-1,到30 h时几乎降解完全.

|
图 8 菌株y5的硝酸盐好氧反硝化过程 Fig. 8 Process of nitrite denitrification of strain y5 |
如图 9所示,氨氮和亚硝态氮共存时,菌体在0-12 h处于生长适应期,从12 h开始进入对数增长期,D600最大为1.11. NH4+-N的浓度由初始的50.62mg ·L-1,经36 h反应后,完全去除. NO2--N的浓度由初始的97.22 mg ·L-1,经36 h反应后,降至19.93 mg ·L-1,去除率为79.50%. NH4+-N从12 h开始快速去除,平均去除速率为1.79 mg ·(L ·h)-1,NO2--N从15 h后开始快速去除,平均去除速率为3.72 mg ·(L ·h)-1. 菌株y5的异养硝化-亚硝酸盐好氧反硝化作用主要发生在对数增长期. 反应过程中几乎没有硝酸盐的产生.

|
图 9 NH4+-N 和 NO2--N混合氮源系统下菌株y5的脱氮过程 Fig. 9 Process of nitrogen removal in the NH4+-N and NO2--N system of strain y5 |
如图 10所示,氨氮和硝态氮共存时,菌体在0-12 h处于适应期,从12 h进入对数增长期,D600最大为1.19. NH4+-N的浓度由初始的51.99 mg ·L-1,经33 h反应后,完全去除,平均去除速率为1.57 mg ·(L ·h)-1. NO3--N的浓度由初始的42.24 mg ·L-1,经30 h反应后,完全去除. NO3--N的还原主要发生在15-21 h,平均去除速率为6.20 mg ·(L ·h)-1. NO2--N在18 h时累积量最大为29.06 mg ·L-1,到27 h时几乎降解完全.

|
图 10 NH4+-N 和 NO3--N混合氮源系统下菌株y5的脱氮过程 Fig. 10 Process of nitrogen removal in the NH4+-N and NO3--N system of strain y5 |
如图 11所示,氨氮、 亚硝态氮和硝态氮共存时,0-9 h菌体处于适应期,9 h进入对数增长期,D600最大为1.10. NH4+-N浓度由53.37 mg ·L-1经36 h反应完全去除,NO3--N浓度由25.19 mg ·L-1经27 h反应完全. NO2--N的浓度由49.26 mg ·L-1,在18 h达到最大值为58.16 mg ·L-1,到27 h反应完全.

|
图 11 NH4+-N、 NO2--N和NO3--N混合氮源系统下菌株y5的脱氮过程 Fig. 11 Process of nitrogen removal in the NH4+-N,NO2--N and NO3--N system of strain y5 |
迄今为止,研究者从土壤、 污水处理反应器、 海洋等环境中分离出的异养硝化-好氧反硝化菌主要包括不动杆菌(Acinetobacter)、 假单胞菌(Pseudomons)、 蜡状芽孢杆菌(Bacillus cereus)及盐单胞菌(Halomonas)等[11]. 本研究从胶州湾深海底泥中分离筛选的菌株y5经鉴定为克雷伯氏菌属(Klebsiella sp.),属于异养硝化-好氧反硝化菌的新菌种,之前关于克雷伯氏菌属异养硝化-好氧反硝化的研究尚没有报道,仅见罗霆等[12]从甘蔗根系分离出1株具有固氮能力的克雷伯氏菌属. 这进一步丰富了异养硝化-好氧反硝化菌的涵盖范围,也为高盐废水的生物处理提供了新的选择.
有机碳源在菌株的HN-AD过程中提供能量和电子,不同碳源因其结构和分子量的差异对菌株的生长和脱氮效果的影响也不同. 赵丹等[13]研究发现菌株ZD8的最佳碳源也为柠檬酸钠. 本实验中,菌株y5在以蔗糖为唯一碳源时,菌株生长最好,但是亚硝酸盐的积累量相当高,以柠檬酸三钠为唯一氮源时,菌株y5生长良好,脱氮效率最高. 综合考虑,菌株y5的最佳碳源为柠檬酸三钠.
反应体系中的pH会直接影响微生物细胞膜的电荷,进而通过影响微生物对营养物质的吸收来影响菌体的生长[14]. 过高或过低的pH均会导致微生物酶的活性降低或丧失[15]. 迄今分离出的异养硝化-好氧反硝化菌大多数适宜pH范围为中性或偏碱. 高喜燕等[16]发现菌株在pH为7.5时的脱氮率最高,但在过酸(pH<5.5)或过碱(pH>9)的条件下,菌株几乎不生长,脱氮率也很低. 黄廷林等[17]筛选出的贫营养菌株A14在pH为7的条件下,对NO3-的去除效果最佳. 在本实验中,菌株y5的最适pH为7.0.
许多研究结果表明,脱氮效率随着C/N的增大而升高,达到一定值后继续增大C/N,脱氮效率不会有太大变化甚至有一定的下降[18, 19]. 本实验也得到类似的结论,当C/N比小于5时,菌株y5的生长量较低,对氨氮和硝态氮的脱除效果也较差. 碳源不足的情况下氨单加氧酶(AMO)和羟氨氧化酶(HAO)对碳源存在竞争,随着C/N的增加,竞争减小,脱氮效率上升. 碳源量增加使细菌的消化性能增强,但过多的碳源会嵌入酶结构中,降低酶的活性,使硝化能力下降[20]. 菌株y5在C/N为9~21时,具有较高的脱氮效率. 最佳C/N为17,C/N继续增高,菌株的生长和脱氮效率降低.
目前人们所认可的氨氮的异养硝化途径主要有两种[21],一种是NH3经AMO氧化为NH2OH,再经HAO氧化为NO2-、 NO3-; 另一种是NH2OH直接转化为N2 O、 N2. 本实验中,以NH4Cl为唯一氮源时,菌株y5的增长速率最大,且反应过程中几乎检测不到NO3--N的存在,判断菌株y5对NH4+-N的异养硝化过程为第二种硝化途径,氨氮经AMO氧化为羟氨后,羟氨在HAO的催化作用下,直接转变为N2 O、 N2从系统中脱除,类似于菌株qy37[22] 的硝化过程. 由于N2 O作为温室气体的辐射增温潜势是CO2的296倍,同时也会破坏大气臭氧层,其存在不可忽视,通过控制工艺的运行参数可以有效地减少N2 O的产生.
以亚硝酸钠和硝酸钾为唯一氮源的条件下,菌株y3均能进行好氧反硝化作用. 以亚硝酸盐为唯一氮源时,菌株的生长速率最小,反应过程中几乎没有硝酸盐的积累,说明亚硝酸盐在亚硝酸盐还原酶的作用下直接还原为氮的气态产物溢出反应系统. 与梁贤等[23]所研究的HN-AD菌YL具有相似亚硝酸盐降解特性. 以硝酸盐为唯一氮源时,检测到大量亚硝酸盐的积累,随后被迅速还原. 表明菌株y5先将NO3--N还原为NO2--N,诱导亚硝酸盐还原酶产生,进而将NO2--N还原为气态氮溢出系统.
氨氮和亚硝酸盐或氨氮和硝酸盐共存时,菌株优先利用NH4+-N进行异养硝化作用,氨氮的降解效率高于以氯化铵为唯一氮源的情况,这表明亚硝酸盐或硝酸盐的加入促进菌体对氨氮的代谢. 原因是亚硝酸盐或硝酸盐诱导反硝化酶的产生,从而加快了硝化-反硝化过程的电子传递速率,促进氨氮的代谢进程. 肖继波等[24]在对Defluvibacter lusatiensis str. DN7的研究中也发现少量的硝酸盐的存在有利于菌株的异养硝化反应,与本实验的结果相类似. 氨氮的存在对菌株y5的好氧反硝化无显著影响,这与Stewart[25]的研究结果一致. 硝酸盐存在时,NO2--N的降解速率显著提高,原因可能是硝态氮的添加,诱导产生硝酸盐还原酶,在促使硝态氮向亚硝态氮转化的同时,刺激亚硝酸盐还原酶的大量产生,进而提高NO2--N的降解速率.
4 结论(1) 从胶州湾海底沉积物中分离筛选出的1株同时具有异养硝化和好氧反硝化能力的菌株y5,经形态观察、 生理生化实验和16S rRNA序列分析,鉴定为克雷伯氏菌属(Klebsiella sp.).
(2) 菌株y5的最佳碳源为柠檬酸三钠,最适pH为7.0,最适C/N为17.
(3) 菌株y5的异养硝化和好氧反硝化作用主要发生在对数增长期,菌株优先利用氨氮进行硝化作用,其次是硝态氮,最后进行亚硝态氮的好氧反硝化. 低浓度的硝酸盐或亚硝酸盐的存在有利于菌株的异养硝化反应,氨氮对好氧反硝化几乎没有影响.
| [1] | Robertson L A, Kuenen J G. Thiosphaera pantotropha gen. nov. sp. nov., a facultatively anaerobic, facultatively autotrophic sulphur bacterium[J]. Journal of General Microbiology, 1983, 129 (9): 2847-2855. |
| [2] | 杨航, 黄钧, 刘博. 异养硝化-好氧反硝化菌Paracoccus pantotrophus ATCC 35512的研究进展[J]. 应用与环境生物学报, 2008, 14 (4): 585-592. |
| [3] | 陈昢圳, 王立刚, 王迎春, 等. 异养硝化-好氧反硝化菌的筛选及脱氮性能的实验研究[J]. 环境科学, 2009, 30 (12): 3614-3618. |
| [4] | 信欣, 姚力, 鲁磊, 等. 耐高氨氮异养硝化-好氧反硝化菌TN-14的鉴定及其脱氮性能[J]. 环境科学, 2014, 35 (10): 3926-3932. |
| [5] | 张培玉, 郭艳丽, 于德爽, 等. 一株轻度嗜盐反硝化细菌的分离鉴定和反硝化特性初探[J]. 微生物学通报, 2009, 36 (4): 581-586. |
| [6] | Wan C L, Yang X, Lee D J, et al. Aerobic denitrification by novel isolated strain using NO2--N as nitrogen source[J]. Bioresource Technology, 2011, 102 (15): 7244-7248. |
| [7] | Yao S, Ni J R, Chen Q, et al. Enrichment and characterization of a bacteria consortium capable of heterotrophic nitrification and aerobic denitrification at low temperature[J]. Bioresource Technology, 2013, 127 : 151-157. |
| [8] | Zhang J B, Wu P X, Hao B, et al. Heterotrophic nitrification and aerobic denitrification by the bacterium Pseudomonas stutzeri YZN-001[J]. Bioresource Technology, 2011, 102 (21): 9866-9869. |
| [9] | 孙雪梅, 李秋芬, 张艳, 等. 一株海水异养硝化-好氧反硝化菌系统发育及脱氮特性[J]. 微生物学报, 2012, 52 (6): 687-695. |
| [10] | Shapovalova A A, Khijniak T V, Tourova T P, et al. Heterotrophic denitrification at extremely high salt and pH by haloalkaliphilic Gammaproteobaeteria from hypersaline soda lakes[J]. Extremophiles, 2008, 12 (5): 619-625. |
| [11] | 苏婉昀, 高俊发, 赵红梅. 异养硝化-好氧反硝化菌的研究进展[J]. 工业水处理, 2013, 33 (12): 1-5. |
| [12] | 罗霆, 欧阳雪庆, 杨丽涛, 等. 1株有固氮能力的甘蔗克雷伯氏菌的分离鉴定及固氮特性[J]. 热带作物学报, 2010, 31 (6): 972-978. |
| [13] | 赵丹, 于德爽, 李津, 等. 菌株ZD8的分离鉴定及其异养硝化和缺氧/好氧反硝化特性研究[J]. 环境科学学报, 2013, 33 (11): 3007-3016. |
| [14] | 王兆阳, 陈国耀, 姜珂, 等. 1株耐冷兼性嗜碱好氧反硝化菌的分离鉴定及反硝化特性[J]. 环境科学, 2014, 35 (6): 2341-2348. |
| [15] | Gupta A B. Thiosphaera pantotropha: a sulphur bacterium capable of simultaneous heterotrophic nitrification and aerobic denitrification[J]. Enzyme and Microbial Technology, 1997, 21 (8): 589-595. |
| [16] | 高喜燕, 刘鹰, 郑海燕, 等. 一株海洋好氧反硝化细菌的鉴定及其好氧反硝化特性[J]. 微生物学报, 2010, 50 (9): 1164-1171. |
| [17] | 黄廷林, 张丽娜, 张海涵, 等. 一株贫营养异养硝化-好氧反硝化菌的筛选及脱氮特性[J]. 生态环境学报, 2015, 24 (1): 113-120. |
| [18] | 李卫芬, 傅罗琴, 邓斌, 等. 1株好氧反硝化菌的分离鉴定及反硝化特性研究[J]. 环境科学, 2011, 32 (8): 2403-2408. |
| [19] | Joo H S, Hirai M, Shoda M. Characteristics of ammonium removal by heterotrophic nitrification-aerobic denitrification by Alcaligenes faecalis No.4[J]. Journal of Bioscience and Bioengineering, 2005, 100 (2): 184-191. |
| [20] | 宋宇杰, 李屹, 刘玉香, 等. 碳源和氮源对异养硝化好氧反硝化菌株Y1脱氮性能的影响[J]. 环境科学学报, 2013, 33 (9): 2491-2497. |
| [21] | Richardson D J, Wehrfritz J M, Keech A, et al. The diversity of redox proteins involved in bacterial heterotrophic nitrification and aerobic denitrification[J]. Biochemical Society Transactions, 1998, 26 (3): 401-408. |
| [22] | 张培玉, 曲洋, 于德爽, 等. 菌株qy37的异养硝化/好氧反硝化机制比较及氨氮加速降解特性研究[J]. 环境科学, 2010, 31 (8): 1819-1826. |
| [23] | 梁贤, 任勇翔, 杨垒, 等. 异养硝化-好氧反硝化菌YL的脱氮特性[J]. 环境科学, 2015, 36 (5): 1749-1755. |
| [24] | 肖继波, 江惠霞, 褚淑祎. 不同氮源下好氧反硝化菌Defluvibacter lusatiensis str.DN7的脱氮特性[J]. 生态学报, 2012, 32 (20): 6463-6470. |
| [25] | Stewart V. Nitrate respiration in relation in to facultative metabolism in enterobacteria[J]. Microbiology Reviews, 1988, 52 (2): 190-232. |