 2015, Vol. 36
2015, Vol. 36



2. 中国科学院大学, 北京 100049;
3. 中国科学院环江喀斯特生态系统观测研究站, 环江 547100
2. University of Chinese Academy of Sciences, Beijing 100049, China;
3. Huanjiang Observation and Research Station for Karst Ecosystems, Chinese Academy of Sciences, Huanjiang 547100, China
土壤微生物量在养分循环与平衡、 土壤理化性质的改善中起着重要作用,指示着生态系统功能的变化[1]. 坡位是重要的地形因子,它可以通过影响土壤微环境、 土壤理化性质、 地上植被的生长及地下碳输入等途径间接影响土壤微生物量[2, 3]. 研究显示,由于长期地表径流侵蚀和淋溶作用,我国大部分地区表层土壤养分和微生物量存在着明显的“洼积效应”,即随着坡位的升高,土壤养分及微生物量会逐渐降低[4, 5, 7],但由于受土地利用方式及干扰程度的影响,上坡位土壤微生物量显著高于下坡位[7]. 但是,对四川寿竹林地的深层土壤的研究结果表明,微生物量对坡位的响应与表层不同[8],这说明不同地质背景土壤的坡位和土层可能对土壤微生物的影响不同. 喀斯特地区土层浅薄、 土壤总量少[9]、 土壤剖面缺乏C层[10]等特点决定其养分及微生物量受土层的影响可能具有自身的独特特征. 土壤微生物直接驱动土壤碳循环过程,且深层土壤碳周转对于碳循环具有重要的作用[11]. 认识喀斯特地区不同坡位深层土壤微生物量,对理解该区深层土壤碳固定及完善土壤碳周转模型具有重要的意义.
目前指征土壤微生物量的测定方法主要包括氯仿熏蒸的土壤微生物生物量、 传统培养的微生物计数、 以及分子生物学的土壤微生物丰度[12, 13]. 已有研究多集中于将土壤微生物生物量碳、 氮来指征土壤微生物量,并重点研究其与土壤肥力以及有关环境因素的关系[14]; 由于土壤中可分离培养的微生物不足1%[15],微生物计数的方法已不能表征土壤微生物量的研究[16],而用土壤微生物丰度指征微生物量的研究少. 细菌、 真菌作为土壤微生物的两大类群,其丰度对外界环境的响应有助于反映土壤微生物质量状况[17]. 喀斯特地区地上和地下二元水土过程的独特性,必然影响土壤养分对坡位与土层的响应具有特殊性,而揭示这种特殊性,是制订喀斯特退化土地植被恢复措施和深入认识土壤碳周转的特征的前提. 以喀斯特峰丛洼地的原生林生态系统为对象,利用氯仿熏蒸法和实时定量PCR技术分别指示土壤微生物生物量与丰度,阐明喀斯特这一独特地质地貌条件下,水土地上与地下二元流失过程是否影响土壤微生物生物量及丰度对坡位和土层的响应,明确以下两个问题: ①坡位和土层对土壤养分和土壤微生物生物量及丰度的影响; ②坡位和土层导致的土壤理化性质变化与土壤微生物生物量及丰度间的关系,旨在为合理利用该区有限的土地资源和制订生态恢复及重建措施提供理论依据.
1 材料与方法 1.1 样地选择及样品采集研究区位于广西环江木论国家自然保护区(107°53′-108°05′ E,25°06′-25°12′ N),属典型喀斯特峰丛洼地景观以及亚热带季风气候区. 海拔400-1000 m,年平均气温15.0-18.7℃,极端高温36℃,极端低温-5℃; 年均降雨量1530-1820 mm,林内相对湿度一般在 80%-90%,夏季可接近饱和; 林区土壤类型简单,主要为石灰土和零星分布的硅质土,均属非地带性土壤; 该区几无人类活动干扰,尤其是没有受到翻耕等农业活动的影响.
于2008年12月至2009年1月,在研究区从山顶到山脚共设置3条样带,分别在每条样带的上、 中、 下坡位各设置一个样方(20 m×30 m),海拔高度分别为615、 509、 416 m. 上、 中、 下坡位共有的优势物种有平阳厚壳桂(Cryptocarya chingii)、 野独活(Miliusa chunii)、 九里香(Murrava exotica)、 茜树(Aidia cochinchinensis); 另外上坡位主要优势物种还包括楠木(Phoebe zhennan)、 灰岩捧柄花(Cleidion bracteosum)等,中坡位为珠子木(Phyllanthodendron anthopotamicum)、 棕木(Cleidion bracteosum)等,下坡位为卵果海桐(Pittosporum ovoideum Gowda)、 山小桔(Glycosmis citrifolia)等,具体的植被多样性信息见表 1. 在样方内采集淋溶层(A 层,0-10 cm)、 过渡层(AB层,30-50 cm)、 淀积层 (B层,70-100 cm)土壤样品. 去除土壤中可见的动植物残体,采用四分法取约50 g左右新鲜土壤置于灭菌自封袋中,封口后立即置于液氮中带回,保存在-80℃冰箱中供土壤微生物丰度分析; 另分取两份: 一份保存在4℃冰箱中供土壤微生物生物量分析,一份经自然风干后用于土壤理化性质分析. 同时,采用环刀法测定土壤容重.
| 表 1 样地植被物种多样性信息 1) Table 1 Diversity index of vegetation in study area |
 ,式中,Pi为第i物种个体数占群落中所有物种个体总数的比例,Pi=Ni/N;H值为Shannon-Wiener指数
,式中,Pi为第i物种个体数占群落中所有物种个体总数的比例,Pi=Ni/N;H值为Shannon-Wiener指数 ;E值为Pielou均匀度指数=H/lnS;不同标识的大写字母代表不同坡位间存在显著差异(P<0.05)
;E值为Pielou均匀度指数=H/lnS;不同标识的大写字母代表不同坡位间存在显著差异(P<0.05)pH采用pH计测定(土水比为1 ∶2.5); 有机质用重铬酸钾容量法-外加热法测定; 全氮用半微量开氏法测定; 全磷用NaOH碱熔-钼锑抗比色法测定; 全钾用NaOH熔融-火焰光度法测定; 碱解氮用碱解扩散法测定; 速效磷用0.5 mmol ·L-1 NaHCO3提取-钼锑抗比色法(Olsen法)测定; 速效钾用NH4OAc浸提-火焰光度法测定[18].
1.3 土壤微生物生物量的测定土壤微生物生物量碳(SMBC)采用氯仿熏蒸浸提-碳分析仪器法,土壤微生物生物量氮(SMBN)采用氯仿熏蒸浸提-全氮测定法测定[12]. 相关计算公式:
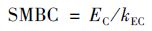
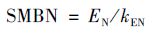
将-80℃冰箱保存的土壤样品置于冷冻干燥器(Freezer Dryer,SIM,USA)抽真空24 h后迅速研磨,用土壤DNA快速提取专用试剂盒(Fast DNA SPIN Kit for Soil,MP)从0.5 g冷冻干燥土壤中提取总DNA,经1.2%琼脂糖凝胶电泳检测总DNA片段大小约为20 kb; 用紫外分光光度计(Nanodrop,PeqLab,Germany)测定总DNA提取量为80.60-584.62 μg ·g-1,A260nm/A280nm比值为1.7-1.9,A260nm/A230nm比值为1.7-1.9,说明所提取的土壤微生物总DNA产量和纯度均较高,可用于进一步的分子生物学分析.
1.5 实时荧光定量PCR扩增采用实时荧光定量PCR(Real-time PCR)测定土壤细菌16S rRNA、 真菌18S rRNA基因拷贝数. 引物分别为:
细菌[19]F1369: 5′-CGGTGAATACGTTCYCGG-3′,R1492: 5′-GGWTACCTTGTTACGACT T-3′; 真菌[20]Fung: 5′-GTAGTCATATGCTTGTCTC-3′,NSI: 5′-ATTCCCCGTTACCCGTTG-3′. 按(Takara)试剂盒的操作说明进行两步法SYBR I 荧光定量PCR扩增(PCR反应液为10 μL体系),定量分析喀斯特峰丛洼地原生林剖面土壤微生物丰度. 实时荧光定量PCR仪: ABI 7900,Foster City,CA,USA.
标准曲线的制作: 分别将细菌、 真菌测序获得的已知种属的阳性克隆子扩增培养后提取质粒DNA,紫外分光光度计检测浓度后,进行10-108倍梯度稀释,作为16S rRNA、 18S rRNA基因的荧光定量的标准样品. 所有样品均4次重复. 以初始模板DNA量C的对数为横坐标,以PCR反应过程中每个稀释样品的Ct值为纵坐标,绘制标准曲线. 细菌和真菌RT-PCR扩增标准曲线分别为:

线性范围分别为104-109 和102-108,扩增效率在0.96-1.09之间,满足基因丰度定量实验要求,且熔解曲线都是单峰型,说明在PCR扩增过程中,没有出现非特异性扩增,由此推断定量PCR扩增所获得的数据是可靠的.
1.6 数据分析以坡位和土层作为处理因子对土壤微生物生物量(SMBC,SMBN)及丰度(细菌、 真菌)进行双因素方差分析,处理间的多重比较用S-N-K法(P<0.05); 坡位和土层导致的土壤理化性质变化与土壤微生物生物量、 丰度间的关系采用Pearson相关性分析及逐步回归分析 (Stepwise regression). 用SPSS 20.0和 Excel 2010做统计分析和制图.
2 结果与分析 2.1 坡位和土层对喀斯特原生林土壤理化性质的影响不同坡位,土壤有机质、 全氮、 全磷、 碱解氮、 速效磷、 速效钾等随土层加深而降低,表现为A层显著高于AB层、 B层(P<0.05); 同一土层,土壤性质对坡位的响应不一,具体表现为: A层、 AB层、 B层土壤下坡位的有机质、 全氮、 全磷、 全钾及碱解氮含量显著高于中坡位、 上坡位(P<0.05),A层土壤的速效钾含量在上坡位表现为最高,而AB层、 B层土壤的速效钾含量则表现为上坡位/下坡位显著高于中坡位(P<0.05); 土壤容重整体表现为下坡位最小; 坡位对pH的剖面分异无显著影响,整体在6.13-7.53范围内(表 2).
| 表 2 土壤理化性质在不同坡位不同土层间的比较 1) Table 2 Comparison on soil physicochemical properties among different slope positions and soil horizons |
 | 图 1 土壤微生物生物量和微生物丰度在坡位与土层间的比较
Fig. 1 Comparison on soil microbial biomass and abundance among different slope positions and soil horizons (a)土壤微生物生物量碳; (b)土壤微生物生物量氮; (c)细菌丰度; (d)真菌丰度 |
土层对土壤微生物生物量的影响显著,同一坡位,A层、 AB层、 B层间存在明显差异,均表现为A层最高(P<0.05)(图 1). 坡位对土壤微生物生物量的影响不一. 土壤微生物生物量碳: A层SMBC在下坡位表现最高,均显著高于中、 上坡位(P<0.05); AB层在中、 上坡位间无显著差异,均高于下坡位; B层则在中坡位最高,上、 下坡位间无显著差异[图 1(a)]. 土壤微生物生物量氮: A层、 AB层、 B层均在下坡位表现为最高,显著高于上坡位(P<0.05)[图 1(b)].
2.3 坡位和土层对喀斯特原生林土壤微生物丰度的影响细菌16S rRNA基因丰度在各坡位各土层的分布(以干土计,下同)为2.61×1010-4.15×1011 copies ·g-1 [图 1(c)]. 其中,同一坡位,沿土层深度的增加,细菌丰度显著减少(P<0.05); A层中坡位/下坡位>上坡位,AB层中坡位>上坡位/下坡位(P<0.05),B层坡位间无显著差异(P>0.05). 真菌18S rRNA基因丰度在各坡位各土层的分布为5.81×106-3.58×108 copies ·g-1[图 1(d)],显著低于细菌丰度. 其中,同一坡位,真菌丰度均表现为A层>AB层>B层; 在3种坡面地形下,A层表现为中坡位>上坡位>下坡位、 AB层无显著差异、 B层表现为中坡位/上坡位>下坡位.
2.4 喀斯特原生林土壤微生物量生物量与丰度的影响因素双因素方差分析结果(表 3)表明,坡位和土层对土壤微生物生物量碳、 氮以及细菌、 真菌丰度具有不同程度的显著影响,同时存在显著的坡位和土壤发生层位交互作用(P<0.01); 从F值来看,土层对土壤微生物量的影响略大于坡位.
相关分析结果表明,除土壤全磷含量与细菌和真菌丰度、 全钾含量与土壤微生物量碳、 微生物量氮和细菌丰度无显著相关性之外,其它土壤性质与土壤微生物生物量与丰度之间均存在显著的正相关关系(真菌丰度分别与全钾显著负相关除外),但值得注意的是,细菌、 真菌丰与速效磷显著正相关(P<0.01),土壤微生物生物量和丰度与速效钾显著正相关(P<0.01).
进一步采用逐步回归分析获得主要因子(表 4),结果表明: 有机质为影响土壤微生物生物量碳的主要因子,偏回归系数为0.632 (P<0.01),随着有机质含量的增加,SMBC也显著增加; 碱解氮为影响土壤微生物生物量氮的正因子,而pH为负影响因子,标准回归系数分别为0.684、 -0.526 (P<0.01); 碱解氮、 速效磷为影响细菌丰度的主要因子,且决定系数R2高达0.718; 速效钾为影响真菌丰度的主要因子 (P<0.05).
| 表 3 坡位及土层对土壤微生物生物量、 丰度的双因素方差分析结果 1) Table 3 Effect of slope position,soil horizon and their interaction on the soil microbial biomass and abundance by two-way ANOVA |
| 表 4 土壤理化性质对微生物量的逐步回归分析 1) Table 4 Key factors among soil physicochemical properties on microbial biomass and abundance by stepwise regression analysis (N=27) |
作为重要的地形因子,坡位可以通过影响土壤的微气候、 土壤的理化性质、 植被的生长以及地下碳输入等途径影响土壤微生物的生长及生物量[2]. 在本研究中,A层土壤微生物生物量碳存在与养分类似的“洼积效应”,均表现为下坡位最高[图 1(a)],与张地等对东灵山栎树[2]、 秦华军等对喀斯特山区寿竹林地[8]的研究一致,表明土壤微生物生物量碳在表层与同一及其他地貌均有相似的变化规律,而AB层,B层则表现为中坡位最高,说明土壤微生物生物量碳是不同坡位上多重影响因素多方面综合作用的结果. 与土壤微生物生物量碳不同,A层、 AB层、 B层的土壤微生物生物量氮均在下坡位最高,显著高于上坡位[图 1(b)],但成毅等[6]在宁夏固原黄土丘陵区天然草地研究结果则显示土壤微生物生物量氮在坡底较低,分析原因可能是由于土壤微生物生物量氮受制于氮素,本研究中全氮、 碱解氮在下坡位显著高于上、 下坡位(表 2),而黄土丘陵区则表现上\中坡位显著高于下坡位[6].
坡位对细菌丰度、 真菌丰度的影响目前研究还较少,结果表明除B层细菌丰度,AB层真菌丰度在各坡位间无显著差异外,均表现为中坡位最高,分析可能的原因: 一方面中坡位既有上坡位冲积物的补充,又有进一步的向下流失[21],使其具有较高的细粒和黏粒含量,加之相对较好的水热条件[22]; 另一方面调查发现中坡位植被草本层Shannon-Wiener指数显著高于上、 下坡位(表 1),而草本层凋落物是土壤异养微生物代谢底物的重要来源,并对土壤微生物数量及组成产生明显的影响[23].
喀斯特区具有独特的水土二元流失特征,其所引发的土壤养分及其驱动的土壤微生物量表现特征与非喀斯特地区既有共性,也具其特殊性. 结果表明,同一坡位,A层,AB层,B层土壤微生物生物量碳、 氮均存在明显差异,表现为A层最高(P<0.05),与前人研究结果[2, 8]一致,说明喀斯特峰丛洼地土壤微生物较为集中的分布于营养状况较好的表层土壤; 同一坡位,细菌、 真菌丰度均随土层深度的加深而显著降低,与Fierer等[24]对加利福尼亚圣塔阳兹山谷的研究结果不同,该研究表明革兰氏阳性菌相对丰度随土层深度的加深而降低,革兰氏阴性菌与真菌则趋势相反,其主要原因可能是喀斯特地区土壤养分的垂直渗漏较非喀斯特区更严重[25](表 2),且底层土壤养分更加贫瘠,因此,底物依赖的异养细菌与真菌丰度显著降低.
土壤微生物生物量指征土壤微生物的生物总质量(以细菌、 真菌为主,还包括放线菌及原生动物等),避免了由于测定土壤微生物的个体和种类数量的技术难度所造成的确定其种群大小的不确定性[16]; 但也忽略了功能各异的微生物所带来的生态效应[26, 27],细菌、 真菌丰度的指征在一定程度上进行了补充与说明. 本研究结果表明,利用土壤微生物生物量及微生物丰度表征土壤微生物量,二者受坡位及土层的影响均比较敏感(表 3),均能作为喀斯特峰丛洼地土壤微环境监测的生物学指标,尽管揭示的主控因子不同(表 4),但可以相互补充. 在坡位和土层的共同影响下,土壤微生物生物量碳的高低主要受土壤中有机碳源的制约,是土壤肥力高低及其变化的重要依据之一[14, 28]; 土壤微生物量氮主要受碱解氮及pH的共同制约,因为凡是影响土壤氮素矿化与固持过程的因素都会影响土壤微生物生物量氮的含量[27],可作为土壤氮素的敏感指示因子. 细菌丰度主要受碱解氮与速效磷的共同制约,真菌丰度则可能受制于速效钾: 喀斯特区一方面全磷的含量并不低(表 2),但是较高pH值和高钙镁含量的土壤具有很强的固磷作用,形成难溶的磷灰石导致磷的有效性降低[28]; 另一方面钾素受成土母质制约较大,且容易随喀斯特地上与地下二元水分流失的影响[5],是主要养分限制因子. 因此可通过细菌丰度初步指征氮、 磷胁迫,真菌丰度指征钾素,仍需通过单因素实验进行下一步的验证.
4 结论(1)坡位和土层显著影响土壤微生物生物量及丰度,且土壤微生物生物量及丰度对土层的响应比坡位相对较强,说明喀斯特地区土壤养分的垂直渗漏相对更应被关注.
(2)由于不同土壤微生物生物量与丰度等微生物指标指示喀斯特土壤养分的状况存在差异,且各指标的受制土壤环境因子不同,在养分胁迫的喀斯特退化生境的生态修复过程中,需要根据具体目标与立地(坡位)条件采用相应策略.
致谢: 感谢中国科学院亚热带农业生态研究所李德军研究员对本文内容的指导.
| [1] | Kuzyakov Y, Subbotina I, Chen H Q, et al.Black carbon decomposition and incorporation into soil microbial biomass estimated by 14C labeling [J].Soil Biology and Biochemistry, 2009, 41 (2): 210-219. |
| [2] | 张地, 张育新, 曲来叶, 等.海拔对辽东栎林地土壤微生物群落的影响[J].应用生态学报, 2012, 23 (8): 2041-2048. |
| [3] | 赵瑜, 钟宇, 张健, 等.不同坡位下巨桉人工林土壤动物群落特征[J].应用生态学报, 2010, 21 (9): 2367-2374. |
| [4] | 高雪松, 邓良基, 张世熔.不同利用方式与坡位土壤物理性质及养分特征分析[J].水土保持学报, 2005, 19 (2): 53-56, 60, 79. |
| [5] | 刘淑娟, 张伟, 王克林, 等.桂西北喀斯特峰丛洼地表层土壤养分时空分异特征[J].生态学报, 2011, 31 (11): 3036-3043. |
| [6] | 成毅, 安韶山, 马云飞.宁南山区不同坡位土壤微生物生物量和酶活性的分布特征[J].水土保持研究, 2010, 17 (5): 148-153. |
| [7] | 邱梅, 张海, 张宇, 等.黄土高原不同坡位苹果林土壤酶活性及微生物量动态变化研究[J].西北农业学报, 2014, 23 (4): 152-159. |
| [8] | 秦华军, 何丙辉, 赵旋池, 等.西南喀斯特山区寿竹林地土壤微生物量与酶活性在不同坡位和剖面层次的分布特征[J].环境科学, 2014, 35 (9): 3580-3586. |
| [9] | 岳跃民, 王克林, 张伟, 等.基于典范对应分析的喀斯特峰丛洼地土壤-环境关系研究[J].环境科学, 2008, 29 (5): 1400-1405. |
| [10] | 朱明秋, 曹建华, 郭芳.基于碳酸盐岩风化的碳源分析及土壤的影响作用机制[J].中国岩溶, 2007, 26 (3): 202-206. |
| [11] | Fontaine S, Barot S, BarréP, et al.Stability of organic carbon in deep soil layers controlled by fresh carbon supply[J].Nature, 2007, 450 (7167): 277-280. |
| [12] | Wu J S, Joergensen R G, Pommerening B, et al.Measurement of soil microbial biomass C by fumigation-extraction-an automated procedure [J].Soil Biology and Biochemistry, 1990, 22 (8): 1167-1169. |
| [13] | Pfaffl M W.A new mathematical model for relative quantification in real-time RT-PCR [J].Nucleic Acids Research, 2001, 29 (9): e45. |
| [14] | 吴金水, 林启美, 黄巧云, 等.土壤微生物生物量测定方法及其应用[M].气象出版社, 2006. |
| [15] | 孙凯, 刘娟, 凌婉娟.土壤微生物量测定方法及其利弊分析[J].土壤通报, 2013, 44 (4): 1010-1016. |
| [16] | 宋长青, 吴金水, 陆雅海, 等.中国土壤微生物学研究十年回顾[J].地球科学进展, 2013, 28 (10): 1087-1105. |
| [17] | Beauregard M S, Hamel C, Atul-Nayyar, et al.Long-term phosphorus fertilization impacts soil fungal and bacterial diversity but not AM fungal community in alfalfa [J].Microbial Ecology, 2010, 59 (2): 379-389. |
| [18] | 鲍士旦.土壤农化分析[M].(第三版).北京: 中国农业出版社, 2008. |
| [19] | Manerkar M A, Seena S, Bärlocher F.Q-RT-PCR for assessing archaea, bacteria, and fungi during leaf decomposition in a stream[J].Microbial Ecology, 2008, 56 (3): 467-473. |
| [20] | Li C G, Li X M, Kong W D, et al.Effect of monoculture soybean on soil microbial community in the Northeast China [J].Plant and Soil, 2010, 330 (1-2): 423-433. |
| [21] | 王恒松, 熊康宁, 刘云.喀斯特区地下水土流失机理研究[J].中国水土保持, 2009, (8): 11-14. |
| [22] | 周萍, 刘国彬, 侯喜禄.黄土丘陵区侵蚀环境不同坡面及坡位土壤理化特征研究[J].水土保持学报, 2008, 22 (1): 7-12. |
| [23] | 蔡锡安, 夏汉平.森林生态系统中草层植物的生态功能[J].热带亚热带植物学报, 2003, 11 (1): 67-74. |
| [24] | Fierer N, Schimel J P, Holden P A.Variations in microbial community composition through two soil depth profiles[J].Soil Biology and Biochemistry, 2003, 35 (1): 167-176. |
| [25] | 陈洪松, 王克林.西南喀斯特山区土壤水分研究[J].农业现代化研究, 2008, 29 (6): 734-738. |
| [26] | Blume E, Bischoff M, Reichert J M, et al.Surface and subsurface microbial biomass, community structure and metabolic activity as a function of soil depth and season[J].Applied Soil Ecology, 2002, 20 (3): 171-181. |
| [27] | 付美云, 杨宁, 杨满元, 等.衡阳紫色土丘陵坡地不同恢复阶段土壤微生物与养分的耦合关系[J].生态环境学报, 2015, 24 (1): 41-48. |
| [28] | Zhang W, Zhao J, Pan F J, et al.Changes in nitrogen and phosphorus limitation during secondary succession in a karst region in southwest China [J].Plant and Soil, 2015, 391 (1-2): 77-91. |