 2015, Vol. 36
2015, Vol. 36



2. 中国地质大学湿地演化与生态恢复湖北省重点实验室, 武汉 430074
2. Hubei Province Key Laboratory of Wetland Evolution and Ecological Restoration, China University of Geosciences, Wuhan 430074, China
据估计,全球土壤总碳库储量约2500 Pg,其中1550 Pg为有机碳库,约为大气中碳储量(约760 Pg)的2倍,为生物碳库(约560 Pg)的2.7倍[1]. 土壤有机碳库的变化将显著影响大气CO2的浓度,因此如何增加土壤有机碳库储量,或是减少土壤有机碳的矿化分解,对缓解全球温室效应具有重要意义.
生物炭(biochar)是黑碳(black carbon)的一种,专指由各种生物质材料(作物秸秆、 木屑、 动物粪便等)在无氧或缺氧条件下,经低温(<700℃)裂解炭化产生的一类高度芳香化的、 抗分解能力极强的固态物质,是一种富含碳元素的有机连续体[2, 3]. 由于生物炭自身碳含量较高并表现出一定的化学或生物学惰性,应用于土壤后其所具有的碳汇功能成为近年来的研究热点[4, 5, 6, 7]. 多数研究者认为生物炭输入土壤后能有效提高土壤总有机碳含量[8, 9, 10],同时对土壤有机碳矿化产生一定影响[11, 12, 13]. 国内外关于生物炭对土壤有机碳矿化影响的研究已经有一些报道,但是结论各不相同,甚至完全相反[14, 15, 16, 17, 18]. 土壤类型不同、 气候条件的差异以及不同的生物炭制备原料和制备条件等因素都会影响生物炭在土壤中的反应性[12, 19],乃至影响其碳汇功能的表现. 到目前为止,当生物炭与土壤混合后,其共存体系中有机碳的分解转化及其环境效应仍然缺乏足够的数据背景作支撑,生物炭输入对土壤本体有机碳矿化的影响与贡献尚不十分明确[20, 21].
本研究以我国长江中下游广泛分布的人为水成土——水稻土为供试土壤(C3土壤),以大宗粮食作物玉米秸秆(C4作物)为制取生物炭的生物质原材料,分别采用水洗(CS)与未水洗(CN)方式处理生物炭,同时模拟自然条件开展室内培养实验,并通过稳定碳同位素技术表征土壤有机碳稳定同位素的变化,以期精确地描述生物炭——土壤系统中生物炭对土壤本体有机碳矿化的影响. 1 材料与方法 1.1 供试材料 1.1.1 生物炭制备及处理
将玉米秸秆洗净,去除表面黏附物,60℃烘至恒重,将秸秆剪碎至1.0 cm的小段,称取20 g已处理好的秸秆于300 mL瓷坩埚中,盖上盖子,置于程序控温马弗炉(KSW-4b-11A,湖北英山县建力电炉制造有限公司)内,以5℃ ·min-1升至目标温度500℃,保持目标温度2 h,关闭马弗炉. 待炉膛温度自然降至室温后,取出烧制好的生物炭,用研钵磨细过100目筛备用.
取部分生物炭与去离子水按1 ∶10比例混合,振荡1h,去除上浮部分生物炭并用砂芯漏斗0.45 μm滤膜过滤,此过程重复5次. 将水洗处理的生物炭置于电热鼓风干燥箱中恒温(25℃)烘干. 水洗处理的生物炭样品记为CS生物炭,未水洗处理的生物炭样品记为CN生物炭.
1.1.2 供试土壤供试土壤采自武汉市江夏区水稻种植区,取样地区连续种植水稻30年以上,为C3土壤. 采用蛇形布点法进行采样,采集0~20 cm的表层土壤,多点混合. 将采集的土壤样品运回实验室、 风干,尽量取出植物残体及其它固体物,磨细过2 mm筛备用. 土壤基本理化性状如下:土壤pH 5.75、 土壤有机质含量19.8 g ·kg-1、 土壤碱解氮含量875 mg ·kg-1、 土壤速效磷含量19.48 mg ·kg-1、 土壤速效钾含量93.60 mg ·kg-1.
1.2 实验方法 1.2.1 室内培养实验实验设置5个处理,每个培养处理土壤质量均为2 kg,生物炭按照不同质量比与土壤均匀混合. 处理1:不添加生物炭的空白对照(CK); 处理2:添加1%水洗生物炭(CS1%); 处理3:添加3%水洗生物炭(CS3%); 处理4:添加1%未水洗生物炭(CN1%); 处理5:添加3%未水洗生物炭(CN3%). 每个处理设置3次重复. 分别将不同用量不同类型的生物炭与土样混合均匀,并调节培养瓶中含水量至田间持水量的 (60±5)%,然后,将盛有20 mL 2mol ·L-1的NaOH溶液的25 mL小烧杯小心地置于培养瓶内,将培养瓶加盖密封好,于(25±1)℃的恒温箱中培养,并设置光照强度(每天 0~12 h: 0 lx; 13~24 h:30000 lx). 模拟自然条件中没有种植植物的土壤. 培养过程中利用重量法定期补充水分.
1.2.2 生物炭与土壤δ13 C值的测定检测方法:同位素质谱分析.
检测仪器:气体稳定同位素比值质谱仪(MAT-251EM,Finnigan MAT,Germany),分辨率:>250(10%谷处); 分辨精度:δ13 C<0.01‰; 半度灵敏:<2×10-6.
有机碳13 C同位素取样测定时间:1、 7、 28、 90、 150 d.
取样方法:将各处理培养土样品风干、 磨细、 过2 mm筛,称取20 g样品加入过量1 mol ·L-1 HCl溶液,充分搅拌,反应24 h以除去碳酸盐类,用去离子水洗涤至中性,室温烘干,过100目筛备用. 测量时取微量已制备好的样品放入气体稳定同位素比值质谱仪中测定,得到δ13 C值.
土壤混合样品碳稳定同位素δ13 C值通过(1)式表达:

生物炭与土壤有机碳对土壤CO2排放量的贡献度可以通过下列公式求得:
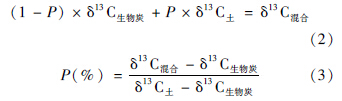
在培养中的第1、 3、 7、 14、 28、 56、 90、 120、 150、 180 d,取出装有碱液的小烧杯,加入过量的BaCl2溶液,加2滴酚酞指示剂,用已标定的盐酸滴定至红色消失,计算CO2的释放量.


采用Microsoft Excel 2007与Croeldraw X4制作数据图、 SPSS 13.0进行单因素方差分析,显著性水平设为0.05. 2 分析与讨论 2.1 生物炭输入对土壤有机碳稳定同位素组成的影响
自然界中C3植物与C4植物具有不同的光合作用路径,在吸收CO2同化形成有机质的过程中,稳定碳同位素分馏模式也不同,导致C3植物与C4植物的δ13 C值差异明显. C3植物的δ13 C 值介于-35‰~-20‰之间,平均为-27‰; C4植物的δ13 C值介于-15‰~-7‰之间,平均为-13‰[24].
C3和C4植物在一个生态系统中长期存在后,就会在土壤有机质中留下同位素信号,土壤有机质源物质主要来自其中所生长的植物,所以土壤有机质的δ13 C值通常接近于该生态系统中优势植物的δ13 C值[22]. 在C3植物长期生长的土壤中,其有机质的δ13 C值因此接近于C3植物的δ13 C值,可以称之为C3土壤.
表 1为水洗生物炭(CS)、 未水洗生物炭(CN)及供试土壤的δ13 C值,2种生物炭的δ13 C值范围在-13.33‰~-13.44‰之间,处于C4植物原有的同位素组成范围内,与C3土壤δ13 C值-27.76‰差异明显.
| 表 1 水洗与未水洗玉米秸秆生物炭及C3土壤的δ13 C值 Table 1 The δ13 C values of washed and unwashed corn straw biochar and C3 soil |
表 2表示通过气体稳定同位素比值质谱仪测定的生物炭——土壤混合样品随培养时间延长,其稳定碳同位素的δ13 C值变化情况.
| 表 2 土壤稳定碳同位素δ13 C随时间的变化值 /‰ Table 2 Changes of the stable isotope of the organic matter in soil,δ13 C value/‰ |
从表 2中发现,随培养时间延长,土壤有机碳同位素的δ13 C值稳定增加,到150 d时,CN3%处理与CS3%处理土壤的δ13 C值分别为-20.415‰和-19.745‰,表示随着C4植物生物炭的加入,土壤有机质δ13 C值是一个稳定增加的过程,说明C4植物秸秆生物炭输入土壤后对土壤有机碳的贡献明显.
2.2 生物炭输入对土壤CO2释放量与释放速率的影响
图 1表示生物炭添加后,不同处理土壤的CO2累计释放量. 从中可知,在180 d的培养期内,CS1%、 CS3%、 CN1%这3种处理的土壤CO2累计释放量分别为1865.7、 1864.4和1856.2 mL ·kg-1,均高于对照土壤CK的1779.0mL ·kg-1,但未形成显著差异. 处理CN3%的CO2累计释放量最大,为2289.1mL ·kg-1,明显高于其他处理及对照土壤. 说明较高添加量下,生物炭经过水洗(CS)后减排效果明显. 这可能是因为水洗处理减少了生物炭中易分解有机碳组分引起的结果,而2种生物炭在较低添加量下,对土壤的CO2累计释放量没有明显影响.
 | 图中误差棒表示标准差(SD),下同 图 1 不同处理土壤CO2释放量累计值 Fig. 1 Accumulative CO2 emissions of different soil treatments |
土壤CO2释放速率是指单位质量土壤单位时间内矿化产生的CO2量,本研究以每千克湿土每天释放的CO2毫升数表示,单位为mL ·(kg ·d)-1.
图 2为不同处理土壤CO2释放速率随培养时间的变化情况,从中可知,培养第1 d,CS1%、 CS3%、 CN1%及CN3%这4种处理CO2释放速率分别为47.0、 57.3、 61.1及74.7mL ·(kg ·d)-1,均显著高于对照土壤42.7 mL ·(kg ·d)-1. 有生物炭添加的土壤矿化CO2释放速率显著增加,其原因可能是供试土壤碳氮(C/N)比值偏低,仅为13.1,在此条件下土壤C成为制约土壤某一类微生物活性的主要因素之一,当生物炭加入后,其较高的含C量与C/N比值使土壤有机C含量增加、 C/N值提高,因此微生物的能量来源增加、 生长环境改善,微生物活性增强,从而导致土壤有机碳矿化速率迅速提高.
 | 图 2 土壤CO2释放速率的动态变化
Fig. 2 Dynamics of soil CO2 release rate
|
对比不同处理生物炭在相同添加量下CO2释放速率发现,CN生物炭处理释放速率明显高于CS处理的CO2释放速率,这一方面可能是由于CN生物炭中易分解组分所引起,另一方面也可能是因为CN生物炭偏碱性、 而CS生物炭更加接近中性,CN生物炭对供试土壤酸性环境的改善更加明显,增强了土壤微生物的酶活性,土壤矿化速率加快.
在整个培养期内,各处理释放速率均随着时间的延长而显著降低,90 d后逐渐稳定,但添加生物炭处理的CO2释放速率依然高于对照土壤.
2.3 生物炭输入对土壤本体有机碳的影响土壤本体CO2释放量等于土壤CO2总排放量乘以土壤本体CO2释放量贡献率,结果如图 3所示. 从中可见,培养150 d后,水洗生物炭处理CS 3%与未水洗生物炭处理CN 3%土壤本体CO2累计释放量分别为1000.4 mL ·kg-1和1153.7 mL ·kg-1,均明显低于对照处理CK的1534.2 mL ·kg-1,说明2种生物炭的添加均能抑制土壤本体有机碳的矿化. 有研究表明添加不同原料、 制备条件的生物炭对土壤本体有机碳矿化均有抑制作用[14, 18],并且随着生物炭添加量增加,土壤本体矿化量下降[17],该结果与本研究结果一致. 但是这种抑制本体有机碳分解的机制并不明确,可能是将微生物利用的碳源转移,使得微生物利用生物炭自身含有的易分解态碳来达到减少对土壤本体有机碳的分解,也有可能是生物炭的添加致微生物的群落发生了变化.
由图 3不难发现,处理CS3%的土壤本体CO2释放量低于处理CN3%的释放量. 通过公式(5)定量计算两种生物炭的激发效应PE值,结果如图 4所示. 从中可知,培养第1 d,CN3%处理有明显的PE正值,CS3%处理土壤本体表现为微弱的PE负值.
 | 图 3 不同处理土壤本体CO2累计释放量
Fig. 3 Accumulative CO2 emissions of the native soil from different samples
|
 | 图 4 生物炭激发效应PE值
Fig. 4 PE value of biochar
|
一般认为外源有机质添加进入土壤后,引起土壤本体有机质矿化速率正的或负的变化是由于微生物活性、 数量和组成的改变引起的[25],可能是因为CN处理生物炭本身含有较多易分解态有机质,提供微生物更多可利用原料,提高了土壤微生物活性,加速了土壤本体有机碳的矿化分解,故短期内引起了正的激发效应.
随着培养时间的延长,CN3%及CS3%处理的PE值均小于0,生物炭添加对土壤的负激发效应迅速增强. 至培养第150 d,处理CS3%的PE值为-34.8%,处理CN3%的PE值为-24.8%. 与未水洗生物炭CN相比,水洗生物炭CS引起的负激发效应更显著. 研究认为,不同生物炭进入土壤后,培养前期将刺激土壤本体有机C的矿化,表现为正激发效应,后期则抑制土壤有机C矿化,产生负激发效应,大部分培养实验持续时间均为半年左右,其总激发效应呈现出不一致的结论[14, 26, 27],其中不同生物炭添加至水稻土中的结果均为负激发效应[17, 18],与本研究结果一致. 3 结论
(1)土壤中添加不同比例的水洗生物炭(CS1%、 CS3%)、 未水洗生物炭(CN1%、 CN3%)后,土壤CO2累计释放量分别为1865.7、 1864.4、 1856.2和2289.1 mL ·kg-1,均高于对照土壤CK的释放量1779.0 mL ·kg-1,对比添加不同处理生物炭土壤的CO2释放量发现,在较低添加量下,不同生物炭对土壤CO2释放量没有明显影响,在较高添加量下,水洗处理生物炭减排效果更显著.
(2)根据对δ13 C值的分析,添加生物炭处理与对照土壤相比,土壤-生物炭混合样品的有机碳稳定同位素δ13 C随培养时间的延长而稳定增加,计算得到水洗(CS3%)与未水洗(CN3%)这2种生物炭处理土壤本体CO2释放量分别为1000.4 mL ·kg-1和1153.7 mL ·kg-1,都显著低于对照土壤的1534.2 mL ·kg-1,这可能是由于生物炭中易分解态碳组分通过代替土壤本体有机碳被微生物利用从而保护土壤本体有机碳免于被分解,产生负激发效应,其中CS生物炭对土壤矿化抑制作用更加明显. 基于180 d的室内培养实验结果,生物炭施用于土壤可作为碳储存载体.
| [1] | Lal R. Soil carbon sequestration impacts on global climate change and food security[J]. Science, 2004, 304 (5677): 1623-1627. |
| [2] | Hammes K, Schmidt M W I, Smernik R J, et al. Comparison of quantification methods to measure fire-derived (black/elemental) carbon in soils and sediments using reference materials from soil, water, sediment and the atmosphere[J]. Global Biogeochemical Cycles, 2007, 21 (3): GB3016. |
| [3] | Goldberg E D. Black carbon in the environment: properties and distribution[M]. New York, NY: John Wiley & Sons, 1985. 198. |
| [4] | Powlson D S, Whitmore A P, Goulding K W T. Soil carbon sequestration to mitigate climate change: a critical re-examination to identify the true and the false[J]. European Journal of Soil Science, 2011, 62 (1): 42-55. |
| [5] | Sutherland W J, Clout M, Cote I M, et al. A horizon scan of global conservation issues for 2010[J]. Trends in Ecology & Evolution, 2010, 25 (1): 1-7. |
| [6] | Woolf D, Amonette J E, Street-Perrott F A, et al. Sustainable biochar to mitigate global climate change[J]. Nature Communications, 2010, 1 (3): 118-124. |
| [7] | Bruun E W, Ambus P, Egsgaard H, et al. Effects of slow and fast pyrolysis biochar on soil C and N turnover dynamics[J]. Soil Biology and Biochemistry, 2012, 46 : 73-79. |
| [8] | 曾爱, 廖允成, 张俊丽, 等. 生物炭对塿土土壤含水量、有机碳及速效养分含量的影响[J]. 农业环境科学学报, 2013, 32 (5): 1009-1015. |
| [9] | Gaskin J W, Steiner C, Harris K, et al. Effect of low-temperature pyrolysis conditions on biochar for agricultural use[J]. Transactions of the Asabe, 2008, 51 (6): 2061-2069. |
| [10] | 章明奎, Bayou W D, 唐红娟. 生物质炭对土壤有机质活性的影响[J]. 水土保持学报, 2012, 26 (2): 127-131. |
| [11] | Hamer U, Marschner B. Priming effects of sugars, amino acids, organic acids and catechol on the mineralization of lignin and peat[J]. Journal of Plant Nutrition and Soil Science, 2002, 165 (3): 261-268. |
| [12] | Spokas K A, Koskinen W C, Baker J M, et al. Impacts of woodchip biochar additions on greenhouse gas production and sorption/degradation of two herbicides in a Minnesota soil[J]. Chemosphere, 2009, 77 (4): 574-581. |
| [13] | 柯跃进, 胡学玉, 易卿, 等. 水稻秸秆生物炭对耕地土壤有机碳及其CO2释放的影响[J]. 环境科学, 2014, 35 (1): 93-99. |
| [14] | 刘燕萍, 高人, 杨玉盛, 等. 黑碳添加对土壤有机碳矿化的影响[J]. 土壤, 2011, 43 (5): 763-768. |
| [15] | Luo Y, Durenkamp M, De Nobili M, et al. Short term soil priming effects and the mineralisation of biochar following its incorporation to soils of different pH[J]. Soil Biology and Biochemistry, 2011, 43 (11): 2304-2314. |
| [16] | Hamer U, Marschner B, Brodowski S, et al. Interactive priming of black carbon and glucose mineralisation[J]. Organic Geochemistry, 2004, 35 (7): 823-830. |
| [17] | 赵次娴, 陈香碧, 黎蕾, 等. 添加蔗渣生物质炭对农田土壤有机碳矿化的影响[J]. 中国农业科学, 2013, 46 (5): 987-994. |
| [18] | 匡崇婷, 江春玉, 李忠佩, 等. 添加生物质炭对红壤水稻土有机碳矿化和微生物生物量的影响[J]. 土壤, 2012, 44 (4): 570-575. |
| [19] | Hilscher A, Heister K, Siewert C, et al. Mineralisation and structural changes during the initial phase of microbial degradation of pyrogenic plant residues in soil[J]. Organic Geochemistry, 2009, 40 (3): 332-342. |
| [20] | Sohi S P, Krull E, Lopez-Capel E, et al. A review of biochar and its use and function in soil[J]. Advances in Agronomy, 2010, 105 : 47-82. |
| [21] | Lehmann J, Sohi S. Comment on "fire-derived charcoal causes loss of forest humus"[J]. Science, 2008, 321(5894): 1295. |
| [22] | 朱书法, 刘丛强, 陶发祥. δ13C方法在土壤有机质研究中的应用[J]. 土壤学报, 2005, 42 (3): 495-503. |
| [23] | Hamer U, Marschner B. Priming effects in different soil types induced by fructose, alanine, oxalic acid and catechol additions[J]. Soil Biology and Biochemistry, 2005, 37 (3): 445-454. |
| [24] | Kuzyakov Y. Sources of CO2 efflux from soil and review of partitioning methods[J]. Soil Biology and Biochemistry, 2006, 38 (3): 425-448. |
| [25] | 黄文昭, 赵秀兰, 朱建国, 等. 土壤碳库激发效应研究[J]. 土壤通报, 2007, 38 (1): 149-154. |
| [26] | Wardle D A, Nilsson M C, Zackrisson O. Fire-derived charcoal causes loss of forest humus[J]. Science, 2008, 320 (5876): 629. |
| [27] | 罗煜, 赵小蓉, 李贵桐, 等. 酸性和碱性土壤中芒草生物质炭激发效应的特征与差异[J]. 土壤学报, 2014, 51 (1): 90-95. |