 2014, Vol. 35
2014, Vol. 35

2. 桂林理工大学环境科学与工程学院, 桂林 541004;
3. 中国地质科学院岩溶地质研究所国土资源部/广西岩溶动力学重点实验室, 桂林 541004;
4. 桂林理工大学广西环境污染控制理论与技术重点实验室, 桂林 541004
 , TANG Hua-feng2, LI Min2, HUANG Bing-fu2, LI Qiang3
, TANG Hua-feng2, LI Min2, HUANG Bing-fu2, LI Qiang3 , ZHANG Jia-yu2, LI Gui-wen2
, ZHANG Jia-yu2, LI Gui-wen2
2. College of Environmental Science and Engineering, Guilin University of Technology, Guilin 541004, China;
3. Karst Dynamics Laboratory, Ministry of Land and Resources of China & Guangxi, Institute of Karst Geology, Chinese Academy of Geological Sciences, Guilin 541004, China;
4. Guangxi Key Laboratory of Environmental Pollution Control Theory and Technology, Guilin University of Technology, Guilin 541004, China
在岩溶生态系统中,土壤有机碳的质量分数要高于非岩溶系统[1,2],因此,研究岩溶生态系统土壤有机碳的积累机制具有重要的意义. 在以水稻土为研究对象所涉及可能的固碳机制中,土壤微生物群落的作用不可忽视[3]. 但是,在岩溶生态系统中,微生物群落是如何调控土壤有机碳循环,人们仍然知之甚少[4]. 因此,以岩溶生态系统为研究对象,探讨微生物群落与土壤有机碳之间的关系可以促进人们对固碳机制的深入理解.
微生物多样性指数和丰度是表征微生物群落特征的两个重要指标. 我国西南岩溶区植被在正向演替的过程中,表层土壤有机碳逐渐积累[5, 6, 7],土壤微生物的组成发生了显著变化[7],可培养微生物及多种功能微生物数量[8]和总的微生物生物量[9,10]均显著提高,土壤微生物的碳源代谢多样性与细菌和真菌的多样性也表现出相同的变化趋势[10,11],这些微生物指标与土壤有机碳质量分数呈显著正相关[10]. 来自不同土地利用方式的研究结果表明,与岩溶区的原生林相比,农田和次生林土壤中微生物的香农-威纳多样指数、 可操作分类单元(operational taxonomic units,OTUs)数量和土壤有机碳质量分数等多个养分指标均有所降低,表明微生物群落的丰度和多样性的降低是加速土壤有机碳损失的重要原因[10,12]. 但是,这两个指标在土壤剖面上的变化及其与土壤有机碳之间的关系还缺乏相应的报道.
位于桂林丫吉村的典型岩溶试验场[13],由洼地、 哑口和坡地这3种不同的地貌组成. 在这个生态系统中,3种地貌土壤剖面层次上可培养微生物丰度的变化规律较为一致,均随着剖面深度的增加而降低,洼地土壤可培养细菌、 真菌和放线菌与多种功能菌群的数量与垭口和坡地也具有明显分异,而且,细菌数量、 微生物数量和土壤酶活性与土壤有机碳质量分数均呈显著正相关关系[14,15]. 但是,土壤微生物多样性的变化以及与土壤有机碳质量分数之间的关系仍不明确. 因此,本文以此岩溶试验场为研究样点,采用聚合酶链式反应-变性梯度凝胶电泳(PCR-DGGE)和荧光定量PCR的方法,分别研究典型岩溶生态系统中细菌和真菌多样性和丰度的变化,探讨二者与土壤有机碳之间的关系,明析微生物多样性和丰度在土壤有机碳积累过程中的作用和意义,以期为深入探讨土壤有机碳的积累机制提供参考依据.
1 材料与方法 1.1 采样点及采样方法桂林丫吉岩溶试验场面积约2 km2,位于广西桂林市东郊8 km的丫吉村,峰丛洼地与峰林平原交界处. 桂林市地处亚热带南端,年平均气温为18.8℃、 年降雨量为1 915.2 mm. 场区的土壤为棕色石灰土,区内土壤覆盖率约为30%,植被覆盖以次生灌丛林为主,覆盖度为60%~80%,表现出喜钙和耐旱性[16,17].
样品采集于2011年11月,位置为1号 洼地、 坡地和垭口. 垭口植被以黄荆为主,土壤厚度30~50 cm,pH 6.00; 坡地植被以黄荆和檵木为主,土壤厚度为70~150 cm,pH 6.55; 洼地植被以老虎刺和构树占优势,土壤厚度为300~500 cm,pH 6.40[14]. 在样地中随机选取3 个近2 m2的采样区,在每个采样区选择3个采样点. 每个采样点分别采集0~10、 10~20和20~30 cm深度的土样,同一采样区3个采样点的土样按层次等比例混合,当天带回实验室. 除去植物残体和入侵体,一部分过2 mm筛,迅速提取土壤微生物DNA以做微生物分析; 另外一部分自然风干,用于土壤理化性质分析. 养分含量见表 1[15].
| 表 1 3种岩溶形态的土壤养分质量分数Table 1 Soil organic carbon,total nitrogen and total phosphorous of three karst forms |
采用Mo BIO公司的DNA快速提取试剂盒(Power Soil)提取土壤微生物总DNA,步骤按操作说明进行. 土壤微生物总DNA用微量紫外分光光度计(Quawell 3000,美国)在260 nm处测定其浓度.
1.2.2 荧光定量PCR用CFX 96(Bio-rad,Hercules,美国)测定16S rRNA和18S rRNA的基因拷贝数. 25 μL PCR反应体系组成如下: 2×SYBR Green real time PCR master mix (Biouniquer,北京) 12.5 μL,去离子无菌水 9.5 μL,DNA模板(genomic DNA,15 ng ·μL-1)1 μL,浓度为20 um ·μL-1正向引物和反向引物各1 μL. 细菌扩增引物采用Muyzer等[18]特异性引物对: 正向F338(5′-ACT CCT ACG GGA GGC AGC AG-3′)和反向R518(5′-ATT ACC GCG GCT GCT GG-3′). 16S rRNA扩增条件为首先是95℃预变性3 min,40个循环为95℃变性1 min,56℃退火1 min,72℃延伸1 min,最后再在72℃延伸 5 min. 以含有16S rRNA V3区片段的质粒作为DNA模板,用从3.42×108~3.42×105系列10倍稀释的梯度的5次重复值画出标准曲线. 从标准曲线得到其扩增效率为87.4%,R2 值为0.962. 18S rRNA扩增引物采用May等[19]的特异性引物对: 正向Fungi(5′-ATT CCC CGT TAC CCG TTG-3′)和反向NS1(5′-GTA GTC ATA TGC TTG TCT C-3′). 18S rRNA扩增引物的反应体系与16S rRNA一致,扩增条件为95℃ 预变性15 min; 40个循环分别为95℃变性1 min,57℃退火1 min,72℃延伸1 min,最后再在72℃延伸5 min,以含有18S rRNA片段的质粒作为DNA模板,用从3.48×107~3.48×103系列10倍稀释的梯度的5次重复值画出标准曲线. 标准曲线上得到18S rRNA的扩增效率为100.9%,R2 值为0.974. 16S rRNA和18S rRNA扩增的溶解曲线中均只有单峰存在,温度分别为89.0℃和90.5℃,说明这两个反应中不存在二聚体污染. 微生物丰度计作每克干土中的基因拷贝数(拷贝 ·g-1).
1.2.3 PCR-DGGE(1)聚合酶链反应(PCR)
PCR用于扩增的引物对和反应体系与1.2.2节中的相同. 有一点不同的是在16S rRNA和18S rRNA正向引物的5′端分别添加40个碱基的GC夹子(5′-CCG CCG CGC GGC GGG CGG GGC GGG GGC ACG GGG-3′). 对细菌而言,PCR反应采用降落(touch down)PCR程序[18],具体如下: 94℃预变性5 min,94℃变性1 min,退火温度从65℃开始,每两个循环降低1℃,一直到55℃,延伸温度为72℃,再在退火温度55℃下作15个循环; 最后在72℃完全延伸5 min. 扩增后的PCR产物用1.2%琼脂糖凝胶电泳检测质量. 对真菌而言,PCR反应条件为: 95℃预变性15 min,91℃变性1 min,57℃退火1 min,72℃延伸2 min,共35个循环; 然后在68℃完全延伸10 min[19]. 扩增后的PCR产物用1.2%琼脂糖凝胶电泳检测质量.
(2)变性梯度凝胶电泳(DGGE)
变性梯度凝胶的制备使用Bio-RAD公司475型梯度制胶系统(Model 475 Gradient Delivery System). 对于细菌,变性梯度从上到下是35%~60%,聚丙烯酰胺凝胶浓度是8%. 在90 V的电压下,60℃电泳9 h; 对于真菌,变性梯度从上到下是15%~30%,在电压100 V下电泳7 h. 电泳完毕后,将凝胶在固定液中固定3 min,用银染液染色10 min,然后用蒸馏水洗涤2次,再加显影液显影3 min; 将染色后的凝胶用Bio-RAD的Gel Doc-2000凝胶影像分析系统拍照; 用Imange lab分析软件(Bio-RAD)确定样品电泳条带的数量、 亮度峰值,用SPSS 13.0进行聚类分析. 多样性指数计算方法参考文献[20]进行,具体如下:
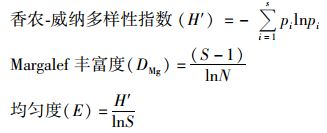
式中,S是OTUs的总数,在DGGE图谱中每一条清晰条带(克隆)被计作一个OTU,N是在每个泳道中所有OUT的总的亮度峰值,pi=ni/N,是第i个OTU在所有克隆中的比值. 1.3 数据处理与统计检验
数据归一化的方法参照文献[21]进行. 首先将所有9个样本的微生物丰度值(或土壤养分)分别进行归一化,得到单个样本归一化的数值,计算公式为; ;然后把每个样本归一化的数值相加后取算术平均值,得到单个样本归一化的数值. 微生物丰度的作图用EXCEL 2003完成. 聚类分析图在SPSS 13.0上完成. 数据间差异的显著性检验以及属性间相关性及其显著性检测均采用JMP 5.0软件进行. 柱状图中的小写字母表示同一岩溶形态不同剖面深度的微生物丰度在P≤0.05水平上显著差异.
;然后把每个样本归一化的数值相加后取算术平均值,得到单个样本归一化的数值. 微生物丰度的作图用EXCEL 2003完成. 聚类分析图在SPSS 13.0上完成. 数据间差异的显著性检验以及属性间相关性及其显著性检测均采用JMP 5.0软件进行. 柱状图中的小写字母表示同一岩溶形态不同剖面深度的微生物丰度在P≤0.05水平上显著差异.
从图 1(a)可以看出,16S rRNA丰度在洼地和垭口土壤剖面随深度的增加总体上呈下降趋势,而在坡地随深度的变化而无显著差异,最高的丰度值出现在洼地土壤的0~10 cm土层,为1.32×1011拷贝 ·g-1. 从图 1(b)可以看出,18S rRNA的丰度值在3种岩溶形态中均随剖面深度的增加而下降,最高的丰度值出现在坡地的0~10 cm土层,为1.16×1010拷贝 ·g-1. 显示16S rRNA和18S rRNA的丰度值随剖面深度的变化规律大致相同. 从表 2可以看出,16S rRNA和18S rRNA总的拷贝数以及二者归一化的拷贝数均随着土壤深度的增加而减小.
 | 图 1 16S rRNA和18S rRNA基因丰度Fig. 1 Abundances of 16S rRNA and 18S rRNA |
| 表 2 岩溶生态系统中微生物的丰度Table 2 Abundances of microbes in karst ecosystem |
从基于16S rRNA和18S rRNA DGGE图谱的分析结果(图 2)可以看出,同一岩溶形态的不同土壤层次均能较好的聚为一类,表明岩溶地貌形态对土壤细菌和真菌多样性的影响较大,这可能与不同岩溶形态土壤的理化性质和植被凋落物差异有关. 从表 3可以看出,16S rRNA的香农-威纳指数和丰富度均随着哑口土壤剖面深度的增加而增大,洼地土壤剖面16S rRNA的香农-威纳指数也有相同的变化趋势. 因此,细菌多样性的变化趋势与细菌的丰度总体上呈现相反的变化趋势. 18S rRNA在哑口土壤的3种多样性指数、 坡地土壤的香农-威纳指数和均匀度与洼地土壤的丰富度均随着剖面深度的增加而增大,表明真菌的多样性与丰度之间呈现出更加明显相反的变化趋势. 微生物丰度与多样性之间的这种关系表明,这两个指标在岩溶生态系统土壤碳循环的指示意义和重要性并不一致.
 | 图 2 16S rRNA和18S rRNA的DGGE图谱和聚类分析Fig. 2 DGGE profiles and their cluster analysis of 16S rRNA and 18S rRNA |
| 表 3 基于16S rRNA和18S rRNA DGGE图谱的微生物多样性指数Table 3 Microbial diversity indices of 16S rRNA and 18S rRNA based DGGE profile |
在岩溶生态系统中,土壤有机碳的质量分数较高[1,2],为微生物的生长提供了可以利用的基质; 而且,较高的有机碳水平改善了土壤的通气状况,提高了水分的固持能力,为微生物的生长提供了适宜的环境条件. 以时间换空间的研究方法表明,岩溶区植被在正向演替的过程中,细菌、 真菌与放线菌及多种参与碳氮循环的功能微生物数量[5]和总的微生物生物量[10,22]均显著提高,土壤微生物的碳源代谢多样与细菌和真菌的多样性指数也表现出相同的变化趋势[10,11],而且,以上微生物指标与土壤有机碳质量分数均呈显著的正相关关系[10, 14]. 来自限制性片段长度多态性的研究表明,与原生林相比,岩溶区农田和次生林的表层土壤中微生物的香农-威纳多样指数、 OTUs数量和土壤有机碳质量分数等多个养分指标均有所降低[23]. 以上这些研究结果表明,在岩溶区土壤剖面的同一层次,微生物的丰度与多样性有着一致的变化趋势. 在岩溶峰丛洼地不同土地利用类型的土壤剖面中,冯书珍等[24]研究也发现,表征微生物特性的多个指标均随着剖面深度的增加而降低,分析植被和坡位是影响表层微生物特性的主要因子. 在本研究所涉及的表层(0~10 cm)土壤中,3种岩溶形态的土壤有机碳质量分数均高于下层,18S rRNA丰度与洼地和垭口16S rRNA的丰度也均显著高于下层土壤,二者总的拷贝数和归一化的拷贝数也高于下层,而且与土壤有机碳、 总氮和总磷的质量分数均有一致的变化趋势,一方面表明这些养分为土壤微生物提供了可供利用的营养,促进了微生物的生长; 另一方面显示这些微生物在表层土壤碳循环和养分循环中具有重要的作用. 而多样性与丰度和土壤养分的变化趋势并不一致,说明多样性除受土壤有机碳和多种养分的影响之外,可能还与不同的岩溶形态、 坡位和地上植被有关,而这些因子对微生物的影响可能是通过凋落物和根系分泌物以及改变土壤的理化因子发挥作用的[24]. 首先,3种岩溶形态土壤的来源不同[25],垭口的土壤为残破积土,坡地的土壤为洪积土,而1号洼地的土壤为冲积土,这种差异可能影响微生物的适应性; 其次,3种岩溶形态中的优势植物不同[14],这可能导致植被通过根系及其分泌物对微生物群落的多样性具有选择作用; 另外,受岩溶形态的影响,土壤水分等理化因子及土壤稳定性[26]也不尽相同,因此也可能造成微生物的多样性差异.
在非岩溶区,洞庭湖典型湿地地区微生物生物量也随着土壤剖面的增加而降低[27]. 来自典型黄土高原不同土地利用类型和不同土壤类型剖面也得出了相似的结论; 而且,微生物生物量均与土壤有机碳呈显著正相关关系,因此,土壤有机碳是影响微生物特性的主要因素[28]. 在本研究所涉及的典型岩溶生态系统中,16S rRNA和18S rRNA的丰度大多随剖面深度的增加而降低.从表 4可以看出,把3种岩溶形态不同土壤剖面微生物丰度作为一个整体,并且结合文献[15]数据分析表明,可培养细菌数量与16S rRNA的丰度呈显著的正相关,可培养真菌数量与18S rRNA的丰度呈显著的正相关,这也进一步证明,微生物丰度指标在土壤养分循环中的重要指示作用. 16S rRNA和18S rRNA的丰度值均与土壤总氮质量分数呈显著的正相关关系(表 4),这可能表明,岩溶土壤的有机碳的质量分数较高,而氮素相对缺乏[29],因此可能成为限制微生物生长的关键因子. 但是,有研究表明,即使在土壤C/N比为28的土壤中,土壤有机碳也是微生物生长的限制因素[30]. 因此,在岩溶生态系统中,土壤碳氮对于维持土壤微生物功能同等重要[31]. 反过来,微生物较高的丰度和功能活性可能加快了碳循环并促进了土壤有机碳积累.
| 表 4 微生物的丰度与可培养微生物数量、 土壤酶活性和养分质量分数的相关性1)Table 4 Correlation between abundance of microbes and culturable microbe number,enzyme activity and nutrient content |
根据本研究中得到的数据,结合文献[15]的数据,把3种岩溶形态的不同剖面得到的数据作为一个整体来做统计分析(表 4),结果表明,在这个典型的岩溶生态系统中,16S rRNA的丰度与脲酶和碱性磷酸酶活性之间均呈极显著的正相关关系,表明细菌(含放线菌)可能在氮素和磷素循环中的作用较大; 18S rRNA的丰度与蔗糖酶活性之间也呈极显著的正相关关系,表明真菌可能是蔗糖酶分泌的主体,因此在土壤碳循环中的具有重要的作用. 16S rRNA和18S rRNA归一化的拷贝数与归一化的土壤酶活性之间呈极显著的正相关关系,这与归一化的可培养微生物数量与土壤酶活性之间显著正相关关系的结论[15]是一致的. 这些相关性数据表明,尽管不同岩溶地貌形态土壤上生长的植被具有差异,但均可能通过凋落物和根系影响土壤微生物的丰度,从而土壤养分尤其是土壤有机碳与微生物的关系最为密切[32]. 因此,较高的土壤有机碳质量分数提高了对微生物的承载能力,多种多样的微生物各自执行特定的功能,互相配合,提高了微生物活性. 因此,在这个典型的岩溶生态系统中,维持一定的微生物丰度是维持土壤碳循环的关键因素.
在本研究中,微生物的多样性指数取决于DGGE图谱中的条带(OTUs)的有无、 多少和丰度值. 分析表明,岩溶区土壤剖面的微生物丰度与多样性呈相反的变化趋势,与土壤酶活性也呈相反的变化趋势,因为绝大多数好氧微生物不能适应养分和氧质量分数较低的环境,而只有少数能够耐受贫瘠和兼性好氧的微生物却能够适应这一环境,因此,微生物丰度降低了,而多样性却增加了. 但是,由于受养分质量分数和氧气体积分数的限制,这种微生物多样性的增加只是意味着少数微生物的种属增加及其数量的少量增加,而相对于庞大的微生物群落来说,增加的数量并不显著,因此总体上微生物数量仍显著降低. 土壤微生物受深层土壤环境的制约[31],生长较为缓慢,活性相对较低,因此在土壤养分循环和碳循环中并不占有主要的地位,表明土壤养分循环不能仅仅依靠少数特定的微生物种群维持,所以维持一定量的微生物数量非常重要. 因此,土壤微生物的丰度(或数量)在土壤养分循环中的意义要大于微生物多样性指数,更适合于作为土壤微生物特性变化的评价指标.
4 结论(1)在典型岩溶地貌形态的土壤中,随着剖面深度的增加,土壤有机碳质量分数显著降低,16S rRNA丰度在洼地和垭口土壤中显著降低,垭口和洼地的香农-威纳指数和垭口丰富度指数也呈降低的趋势; 18S rRNA的丰度也显著降低,大多数多样性指数也呈下降的趋势.
(2)土壤有机碳和微生物丰度与多样性指数之间总体上呈相反的变化趋势,表明土壤微生物丰度指标在岩溶土壤碳循环中的意义要重于微生物多样性指数.
| [1] | Ahmed Y A R, Pichler V, Homolák M, et al. High organic carbon stock in a karstic soil of the Middle-European Forest Province persists after centuries-long agroforestry management [J]. European Journal of Forest Research, 2012, 131 (6): 1669-1680. |
| [2] | 曹建华, 袁道先, 潘根兴. 岩溶生态系统中的土壤[J]. 地球科学进展, 2003, 18 (1): 37-44. |
| [3] | 潘根兴, 李恋卿, 郑聚锋, 等. 土壤碳循环研究及中国稻田土壤固碳研究的进展与问题[J]. 土壤学报, 2008, 45 (5): 901-914. |
| [4] | Chapin Ⅲ F S, McFarland J, David McGuire A, et al. The changing global carbon cycle: linking plant-soil carbon dynamics to global consequences [J]. Journal of Ecology, 2009, 97 (5): 840-850. |
| [5] | 周玮, 周运超. 北盘江喀斯特峡谷区不同植被类型的土壤酶活性[J]. 林业科学, 2010, 46 (1): 136-141. |
| [6] | 胡忠良, 潘根兴, 李恋卿, 等. 贵州喀斯特山区不同植被下土壤C、N、P含量和空间异质性[J]. 生态学报, 2009, 29 (8): 4187-4195. |
| [7] | Li L Q, Wang D, Liu X Y, et al. Soil organic carbon fractions and microbial community and functions under changes in vegetation: a case of vegetation succession in karst forest [J]. Environmental Earth Sciences, 2014, 71 (8): 3727-3725. |
| [8] | 龙健, 李娟, 滕应, 等. 贵州高原喀斯特环境退化过程土壤质量的生物学特性研究[J]. 水土保持学报, 2003, 17 (2): 47-50. |
| [9] | 张平究, 潘根兴. 不同植被群落下喀斯特土壤养分及生物化学性质特征[J]. 水土保持学报, 2012, 26 (1): 77-80, 84. |
| [10] | Zhu H H, He X Y, Wang K L, et al. Interactions of vegetation succession, soil bio-chemical properties and microbial communities in a Karst ecosystem [J]. European Journal of Soil Biology, 2012, 51 : 1-7. |
| [11] | He X Y, Wang K L, Zhang W, et al. Positive correlation between soil bacterial metabolic and plant species diversity and bacterial and fungal diversity in a vegetation succession on Karst [J]. Plant and Soil, 2008, 307 (1-2): 123-134. |
| [12] | Chen X B, Su Y R, He X Y, et al. Soil bacterial community composition and diversity respond to cultivation in Karst ecosystems [J]. World Journal of Microbiology and Biotechnology, 2012, 28 (1): 205-213. |
| [13] | 袁道先. 全球岩溶生态系统对比: 科学目标和执行计划[J]. 地球科学进展, 2001, 16 (4): 461-466. |
| [14] | 李为, 余龙江, 李涛, 等. 岩溶生态系统土壤酶活性的时空动态及其与土壤肥力的关系——以桂林丫吉村岩溶试验场为例[J]. 农业环境科学学报, 2008, 27 (1): 260-266. |
| [15] | 靳振江, 李强, 黄静云, 等. 典型岩溶生态系统土壤酶活性、微生物数量、有机碳含量及其相关性——以丫吉岩溶试验场为例[J]. 农业环境科学学报, 2013, 32 (2): 307-313. |
| [16] | 刘再华, 何师意, 袁道先, 等. 土壤中的CO2及其对岩溶作用的驱动[J]. 水文地质工程地质, 1998, 25 (4): 42-45. |
| [17] | 潘根兴, 曹建华, 何师意, 等. 土壤碳作为湿润亚热带表层岩溶作用的动力机制: 系统碳库及碳转移特征[J]. 南京农业大学学报, 1999, 22 (3): 49-52. |
| [18] | Muyzer G, De Waal E C, Uitterlinden A G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA [J]. Applied and Environmental Microbiology, 1993, 59 (3): 695-700. |
| [19] | May L A, Smiley B, Schmidt M G. Comparative denaturing gradient gel electrophoresis analysis of fungal communities associated with whole plant corn silage [J]. Canadian Journal of Microbiology, 2001, 47 (9): 829-841. |
| [20] | Hill T C J, Walsh K A, Harris J A, et al. Using ecological diversity measures with bacterial communities [J]. FEMS Microbiology Ecology, 2003, 43 (1): 1-11. |
| [21] | 靳振江, 邰继承, 潘根兴, 等. 荆江地区湿地与稻田有机碳、微生物多样性及土壤酶活性的比较[J]. 中国农业科学, 2012, 45 (18): 3773-3781. |
| [22] | 兰雪, 戴全厚, 喻理飞, 等. 喀斯特退化森林不同恢复阶段土壤酶活性研究[J]. 农业现代化研究, 2009, 30 (5): 620-624. |
| [23] | 陈香碧, 苏以荣, 何寻阳, 等. 不同干扰方式对喀斯特生态系统土壤细菌优势类群——变形菌群落的影响[J]. 土壤学报, 2012, 49 (2): 354-363. |
| [24] | 冯书珍, 苏以荣, 秦新民, 等. 喀斯特峰丛洼地土壤剖面微生物特性对植被和坡位的响应[J]. 生态学报, 2013, 33 (10): 3148-3157. |
| [25] |
袁道先, 戴爱德, 蔡五田, 等. 中国南方裸露型岩溶峰丛山区岩溶水系统及其数学模型的研究[M]. 桂林: 广西师范大学出版社, 1996. 1-4. [26] 李强, 靳振江, 李忠义, 等. 岩溶地貌部位对土壤酶活性和微生物丰度的影响[J]. 水土保持通报, 2014, 34 (3): 50-54. |
| [26] | 彭佩钦, 张文菊, 童成立, 等. 洞庭湖典型湿地土壤碳、氮和微生物碳、氮及其垂直分布[J]. 水土保持学报, 2005, 19 (1): 49-53. |
| [27] | 党亚爱, 李世清, 王国栋, 等. 黄土高原典型土壤全氮和微生物氮剖面分布特征研究[J]. 植物营养与肥料学报, 2007, 13 (6): 1020-1027. |
| [28] | 闫俊华, 周传艳, 文安邦, 等. 贵州喀斯特石漠化过程中的土壤有机碳与容重关系[J]. 热带亚热带植物学报, 2011, 19 (3): 273-278. |
| [29] | Demoling F, Figueroa D, Bth E. Comparison of factors limiting bacterial growth in different soils [J]. Soil Biology and Biochemistry, 2007, 39 (10): 2485-2495. |
| [30] | 宋敏, 邹冬生, 杜虎, 等. 不同土地利用方式下喀斯特峰丛洼地土壤微生物群落特征[J]. 应用生态学报, 2013, 24 (9): 2471-2478. |
| [31] | Cavagnaro T R, Jackson L E, Scow K M, et al. Effects of arbuscular mycorrhizas on ammonia oxidizing bacteria in an organic farm soil [J]. Microbial Ecology, 2007, 54 (4): 618-626. |