 2014, Vol. 35
2014, Vol. 35



2. 中国科学院生态环境研究中心城市与区域生态国家重点实验室, 北京 100085
2. State Key Laboratory of Urban and Regional Ecology, Research Center for Eco-Environmental Sciences, Chinese Academy of Sciences, Beijing 100085, China
目前的O3浓度相对于工业化前已经增加了36%[1]. 随着我国工业化和城市化进程的加快,能源消耗不断增加,大气污染危害日益严重,尤其是近地层O3浓度的升高,导致农作物产量降低,森林衰退[2, 3, 4, 5, 6]. Ding 等[7]通过对 MOZAIC (measurement of ozone and water vapor by airbus in-service aircraft) 航测数据的研究发现,在过去10年里我国东部地区地面臭氧浓度以每年 2%左右的速度增长. Wang等[8]利用 1994~2007 年 14年间香港地区监测数据得出,中国南方地区臭氧的背景值浓度每年增加 0.58 nL ·L-1. 在我国,随着社会经济的快速发展,无论珠江和长江三角洲,还是京津地区,空气O3浓度一直较高.
有关臭氧对植物的影响研究多集中在地上部分,如臭氧降低气孔导度、 抑制光合作用、 产生叶面伤害、 降低植物生物量、 作物产量及森林生态系统生产力等[9, 10, 11, 12, 13],而关于臭氧对土壤微生物的影响研究还比较少[14]. 但是臭氧能改变植物光合产物分配[15],降低植物对根系的碳分配,而转移到根系的光合产物减少也会影响根系微生物之间的相互作用[16, 17],同时臭氧浓度提高会降低凋落物的质量和分解速率,从而降低土壤微生物生物量,并且改变微生物的群落结构及功能多样性[18]. 臭氧浓度升高抑制小麦根系土壤微生物的活性,降低根系土壤微生物代谢多样性[19, 20]; 臭氧也降低水稻土壤微生物的代谢功能,同时改变了土壤微生物的磷脂脂肪酸(PLFA)组成[21],但臭氧对土壤微生物群落结构的具体影响不明确,如臭氧主要影响的是土壤微生物中哪些磷脂脂肪酸等.
随着土壤微生物研究越来越广泛、 深入,有很多方法被引入到土壤微生物分析中,其中,PLFA生物标记法对实验条件要求较低,方法灵敏[22],并能够快速、 直接、 有效地提供较为客观可靠的土壤微生物含量、 组成及群落信息[23,24]等优点被广泛运用于土壤微生物分析中. 稳定同位素标记技术作为一种快速发展的新技术,已经被应用于土壤微生物学研究. PLFAs技术与稳定同位素分析的结合,可以很好地识别复杂土壤生态系统中利用外加底物的微生物种群,确定生态系统在外来胁迫因素作用下,不同类群土壤微生物参与有机质代谢的功能. 采用稳定性同位素标记技术,通过直接测定不同土壤微生物PLFA类群的稳定性同位素丰度变化及其对臭氧熏蒸的响应,以期回答:①植物同化的稳定性同位素标记过的CO2能否很快传递到土壤微生物中?②传递到土壤微生物中的同位素丰度是否受到臭氧熏蒸的影响?③如果是,哪些微生物类群将受到何种影响?本研究利用13 C同位素标记和PLFA技术,对臭氧浓度升高后土壤微生物结构的响应过程进行探讨,以明确臭氧改变光合产物分配引起的土壤微生物的变化,以期为了解未来近地表大气臭氧浓度持续增加情况下生态系统地下过程的响应规律提供基础数据和理论依据.
1 材料与方法 1.1 室内熏气系统室内实验所用的熏气装置采用专门定做的研究大气污染对植物影响的柜式气室(1.2 m×1.2 m×2.1 m)[25]. O3由臭氧发生器产生,以纯氧作为O3发生器的气源,O3由箱底部进入生长箱,进入前先与鼓风机吹进的空气混合,再经过箱子中下部起混合作用的薄钢板(孔径1 mm),使O3与空气混合均匀. 箱子中部为带孔钢板(孔径1 mm),供放盆钵和再次混匀气体. 充分混匀后的气体进入生长室,生长室内O3浓度由O3分析仪监测 (ML98l0型,ECOTECH公司),O3浓度通过空气钢瓶供气量的大小和质量流量计控制. 生长箱顶部有排气通道,用于熏蒸后废O3气体的排放.
1.2 O3的暴露处理水稻暴露实验采用了两个处理,即对照CK和臭氧浓度升高的处理O3. CK为空气对照(直接向气室内通空气,其臭氧浓度约4~10 nL ·L-1). 现有的研究大多是在恒定浓度下进行臭氧熏蒸,这并不能完全真实的反应实际情况,实际上大气环境中的臭氧会有一个明显的日变化,从清晨开始臭氧浓度会随光照等条件有所升高,14:00~15:00臭氧浓度达到一天的最高值,之后臭氧浓度又会缓慢回落. 为了真实反映大气中03浓度变化,本研究采用具有日变化的臭氧熏蒸方式,臭氧浓度升高处理的配气方案为09:00时臭氧浓度约40 nL ·L-1,每过0.5 h臭氧浓度升高20 nL ·L-1,直到12:30达到一天的最大值为180 nL ·L-1持续1 h,随后臭氧浓度与上午呈对称式下降,到17:00回落到40 nL ·L-1,最大值为170 nL ·L-1,其日平均为110 nL ·L-1.
1.3 水稻的种植管理水稻品种为3694繁,采用的土是从浙江嘉兴野外实验基地采回的田间土,过2mm筛. 移栽水稻前在土壤中添加一定的肥料,0.428 g ·kg-1尿素、 0.323 g ·kg-1 CaHPO4 ·2H2O、 0.247 g ·kg-1 K2SO4以及0.4732 g ·kg-1 Ca(H2PO4)2 ·H2O. 水稻种子于6月5日用5% H2O2 消毒后置于培养皿中于25℃培养箱中催芽3 d,然后在珍珠岩苗床上培养,培养期间用Hoagland营养液浇灌,7月2日将水稻苗从珍珠岩上移植到高30 cm、 直径15 cm的PVC筒中,每个臭氧处理下15盆,每盆2株,于室外自然光照下生长. 7月20日将栽有水稻苗的PVC筒移入气室内,开始熏气,11月2日熏气结束. 该气室内白天温度为25~30℃,夜间为20~25℃; 相对湿度为50%~85%,CO2浓度与正常大气中一致,光源是波长400~700 nm、 光强220 μmol ·(m2 ·s)-1的人工光源的,光照时间是07:00~19:00. 生育期间每天转换PVC筒的方向以保证水稻光照均匀.
1.4 13 C同位素示踪分别于8月21日和9月19日进行两次13 CO2标记,标记实验在自制的标记室内进行. 标记装置由一个水封槽底座有机玻璃罩子组成,底座上装有一个空气压缩机,温度控制器. 由于标记时日照很强,会使有机玻璃标记室内温度过高产生雾气,因此利用空气压缩机进行冷却,同时利用温度控制器将标记室内温度稳定在一个温度范围内从而避免雾气产生影响植物的光合作用. 将栽有水稻植株的PVC筒放在水封槽内的底座上,每盆水稻在PVC筒口平面上用黑色塑料布遮住(水稻正常露出),以避免盆内水体中的藻类等进行光合作用吸收13 CO2. 每次标记时两个处理各标记3盆,共在标记室内放6盆水稻. 水稻放好后将有机玻璃罩子罩在水封槽上,保证其密封性,13 CO2气体由箱体上方的孔注入. 同时用CO2红外分析仪监测标记室内的CO2 浓度.
植株放入后,先将室内气体通过一个过滤器进行循环以吸收掉背景的CO2,使背景CO2浓度降至100×10-6左右,将此通路关闭,再开始注入13 CO2. 13 CO2由Ba13 CO3(98% atom%)与磷酸反应产生,产生的气体用气袋收集,然后用注射器分次向气室内注入该气体. 标记从09:00开始向气室内注入13 CO2气体,使气室内的13 CO2浓度维持在250×10-6左右,共标记6 h,之后再注入普通CO2气体使室内残留的13 CO2能完全吸收. 两次标记分别消耗Ba13 CO3约10 g左右,且第一次标记时植株的光合能力要强于第二次标记的时候. 未标记的样品则用CO2 代替13 CO2进行上述过程.
1.5 样品采集标记完成后的植株从标记室内取出后继续放回到相应处理的气室内继续进行臭氧暴露处理,标记72 h后取根际土壤样品,分别进行微生物量测定(包括13 C-微生物量碳)和磷脂脂肪酸(13 C-PLFA)分析.
1.6 分析测定 1.6.1 土壤样品中13 C-微生物量的分析测定土壤样品采样后按照微生物碳的测定方法进行熏蒸,用TOC分析仪测定出熏蒸和未熏蒸土壤样品的有机碳,计算其微生物量碳. 然后分别取20 mL熏蒸和未熏蒸样品提取液在80℃烘干并磨成粉末,用同位素比率质谱仪测定13 C. 13 C的原子百分比计算公式为[23]:


13 C-PLFA按照上述PLFA同样的方法提取后,用GC-c-IRMS进行PLFA分析及每个单峰PLFA的δ13 C的确定.
PLFA单体的量按照公式(5)进行计算:

每个单体PLFA中的13 C的量按照标记样品和未标记样品的差值进行计算:
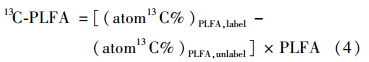
单体PLFA中13 C的相对丰度(the relative abundance) 按公式(5)进行计算:

第一次取样时臭氧对土壤微生物量碳没有影响,但后两次取样臭氧则显著降低了水稻土壤的微生物量碳,相对于空气对照而言臭氧处理后水稻土壤微生物量碳分别降低 26.1%和14.6%. 但第一、 二次标记后的臭氧处理的土壤样品中微生物量的 13 C均是显著降低的,分别比空气对照降低 了68.6%和59.1% (表 1).
| 表 1 臭氧对水稻土壤微生物量碳及13 C的影响1)Table 1 Effects of ozone on soil microbial biomass carbon and 13C partition under rice |
没有13 CO2 标记的样品中无论是熏蒸还是未熏蒸的土壤提取液中的δ13 C(‰)值很接近,随时间也没有发生变化,未熏蒸土壤提取液的δ13 C(‰)为-24.06‰±0.17‰,熏蒸土壤提取液中δ13 C(‰) 为-25.36‰±0.03‰. 13 CO2标记的样品中,熏蒸土壤提取液的δ13 C(‰)明显的高于未熏蒸土壤提取液,且9月22日样品中熏蒸土壤提取液的δ13 C(‰)比未熏蒸土壤提取液增加得要比8月24日多. 随时间的延长在9月22日样品中无论是熏蒸还是未熏蒸土壤提取液中的δ13 C(‰)都要相应地高于8月24日的样品.
对于未熏蒸样品而言,CK 和O3处理之间δ13 C(‰)没有差别; 而对于熏蒸样品而言,8月24日对照处理下δ13 C(‰)为-6.69‰,O3处理下δ13 C(‰)为-8.44‰,9月22日对照处理和O3处理下δ13 C(‰) 分别为0.49‰和-0.60‰,O3处理明显降低了熏蒸土壤提取液的δ13 C(‰).
 | CKUL,对照处理中未标记13 CO2的样品; O3UL,O3处理中未标记13 CO2样品; CKL,对照处理中标记13 CO2的样品; O3L,O3处理中标记13 CO2的样品图 1 13 CO2标记与未标记两个处理中熏蒸土壤样品和未熏蒸土壤样品的δ13 CFig. 1 δ13 C of fumigated and unfumigated soil extracts of soil samples from pots with and without 13 CO2 labelling of rice plants under control and O3 treatments |
室内水稻土壤中一共检测到11种C14~18的脂肪酸,包括直链饱和脂肪酸、 支链饱和脂肪酸、 环丙基脂肪酸以及多不饱和脂肪酸,其中含量最丰富的脂肪酸是16:0、 15:0、 a15:0、 i16:0以及cy17:0,均占总量的10%左右.
第一次标记时水稻土壤PLFA主成分分析表明,主成分1和2分别解释了变异量的69.1%和23.3%,但并没有区分开臭氧处理和对照处理,说明第一次标记时两个处理间PLFA结构没有差异[图 2(a)].
 | 图 2 两次标记时水稻土壤微生物PLFA结构Fig. 2 Plot of Factor 1 versus Factor 2 scores from principal components analysis of PLFA data between rhizosphere soils labeling on 2424 Aug. and 19th Sep. |
第二次标记时水稻土壤PLFA主成分分析表明,主成分1和2分别解释了变异量的57.1%和28.8%,且得分系数方差分析表明PC1明显将CK和O3处理区分开来(F=22.1,P=0.009,PC1),说明第二次标记时臭氧已经明显的改变了水稻土壤PLFA的结构. 造成PC1显著分异的脂肪酸是与其显著正相关的14:0和cy17以及与PC1显著负相关的i16:0和18:2ω6,9这4种脂肪酸,且臭氧处理后前两种脂肪酸的含量是降低的,而后两种脂肪酸则是增加的[图 2(b)].
水稻植株进行13 CO2标记后,部分13 C迅速转移到土壤中,进入微生物的PLFA中,但13 C并不是均匀分布于PLFAs中. 两次标记前后δ13 C值增加最多的均是14:0和cy17以及15:0,其余脂肪酸的δ13 C值增加幅度差不多. 第一次标记结果表明,臭氧熏蒸显著的改变了除17:0以外其他PLFA单体的δ13 C值,i16:0和i17:0这2种脂肪酸在臭氧处理后δ13 C值显著提高,而其他6种细菌脂肪酸包括14:0、 15:0、 a15:0、 16:0、 cy17:0、 18:0,以及放线菌指示脂肪酸18:0(10Me)和真菌脂肪酸18:2ω6的δ13 C值均显著降低 [图 3(a)]. 而第二次标记后,则臭氧对PLFA单体δ13 C值的影响没有第一次明显,i16:0经臭氧处理后δ13 C值仍显著增加,同时臭氧显著降低了14:0、 15:0、 16:0、 a15:0、 i17:0、 cy17这6种脂肪酸的δ13 C值,而对放线菌的指示脂肪酸18:0(10Me)以及真菌脂肪酸18:2ω6,9的δ13 C值都没有产生影响[图 3(b)].
 | 图 3 两次标记后土壤样品13 C-PLFA单体的Δδ13 C ‰值Fig. 3 Δδ13 C ‰ of individual 13 C-PLFA of labeled and unlabeled soils under different O3 treatments on 24th Aug. and 22nd Sep.*、 **、 ***分别表示CK处理和O3处理在0.05、 0.01、 0.001水平上差异显著,下同 |
PLFA单体的13 C相对丰度(13 C%)表明,16:0和cy17这2种脂肪酸占有PLFA中总的 13C的一大半,两者总的13 C%超过了60%,其次13 C%含量高的脂肪酸包括14:0、 15:0、 a15:0和i16:0. PLFAs单体中13 C%的差别表明微生物群落对输入的光合产物存在不同的反应,臭氧作用后会影响不同PLFAs单体中的13 C%. 第一次标记样品中,相对于空气对照土壤而言,臭氧显著提高了4种饱和脂肪酸的13 C%,包括15:0、 16:0、 17:0以及a15:0; 同时显著降低了i16:0和i17:0这2种脂肪酸的13 C相对含量,而对放线菌脂肪酸和真菌脂肪酸中13 C相对含量没有影响[图 4(a)]. 第二次标记样品中,臭氧显著提高了两种细菌脂肪酸16:0、 17:0,放线菌脂肪酸18:0(10Me)以及真菌脂肪酸18:2ω6的13 C的分配量(13 C%); 同时显著抑制了13 C对14:0和cy17这2种脂肪酸的分配[图 4(b)].
 | 图 4 两次标记后土壤样品中PLFA单体的13 C含量Fig. 4 13 C incorporation into phospholipid fatty acids (PLFAs) for the fourth labeling event performed on 24th Aug. and 22nd Sep. |
由于臭氧作用于植物叶片导致光合作用降低,总的光合产物降低,但植物本身存在一个自我修护机制,会用更多的碳来修补叶片的损伤,从而导致输入到茎和根部的碳减少[27]. 本研究也证明了这一机制的存在,臭氧胁迫后植物叶片碳分配增加而茎和根等其他部位碳分配减少[13]. 这一结果也与Ariyaphanphitak等[28]得到的研究结果一致. 臭氧胁迫后对根部碳分配的降低会减少根系分泌物的量,从而导致土壤微生物可利用的碳源减少,势必会影响到土壤微生物的量. 本研究表明,虽然第一次标记时臭氧胁迫下的土壤微生物总的生物量碳没有发生变化,但13 C-MBC已经明显降低,第二次标记时则总的生物量碳和13 C-MBC均受到臭氧的明显抑制,这说明臭氧明显的降低了即时光合产物进入到土壤微生物中的输入量. 另外的研究发现,臭氧浓度升高降低小麦土壤微生物生物量碳[19, 20]; OTC实验表明臭氧浓度升高同样降低了大田水稻土壤微生物生物量碳[21]. 关于臭氧对土壤微生物数量和生物量的影响,不同研究结果也不尽一致. Larson等[29]研究发现当平均臭氧浓度为54 nL ·L- 1 时,暴露于其中的温带森林的土壤微生物碳(95 μg ·g-1)要比环境臭氧浓度下的土壤微生物碳高(70 μg ·g-1),但二者之间差异不显著,不同臭氧处理之间的土壤微生物呼吸也没有显著差异. Shafer[30] 研究臭氧和酸雨对高粱根际微生物的影响,发现根际细菌的总数与臭氧呈现一个曲线关系,臭氧和酸雨趋向于刺激根际真菌繁殖体的数量,但这种影响不显著. 当酸雨pH 4.0时,根际具磷酸酶活性的细菌数量随臭氧呈线性增加. 根际的其它微生物与臭氧和酸雨的相互作用没有很大关系.
本研究结果表明,水稻植株进行13 CO2标记后,少部分光合产物迅速转移到土壤中,成为微生物PLFA组成部分,14:0、 16:0和cy17这3种脂肪酸是13 C丰度最高,也是13 C分配最多的脂肪酸. 第一次标记后,臭氧显著影响13 C分配的脂肪酸主要是细菌的脂肪酸,而对放线菌和真菌脂肪酸的13 C分配没有影响; 而到了第二次标记的时候则臭氧影响13 C分配的细菌脂肪酸减少,但显著提高了放线菌和真菌脂肪酸中的13 C的分配. 这可能是由于在前期细菌生长迅速且能分解水稻分泌的一些简单化合物,从而对臭氧胁迫产生的碳源改变的反应比较敏感; 而相对于细菌而言,放线菌和真菌是分解一些较难分解的大分子化合物,由于它们生长相对缓慢,前期标记时生物量还相对少,因此对由于臭氧胁迫引起的碳源变化反应不敏感. 但到了后期,随着简单化合物的分解其量在减少,可供细菌利用的碳源减少,从而受到碳源影响的PLFA种类也减少,即臭氧影响碳分配的PLFA种类减少; 而随着生长繁殖的进行放线菌和真菌的生物量增加,同时难分解化合物的累积为放线菌和真菌提供了丰富的碳源,此时臭氧胁迫引起的碳源变化就会对放线菌和真菌产生明显的影响,表现为臭氧胁迫下放线菌和真菌13 C分配增加. Manning等[31]以豆类(Pinto bean)为研究对象,发现暴露在臭氧中的豆根比过滤空气中的豆根能分离出更多的真菌,经臭氧处理与未经臭氧处理的豆根被相同真菌定殖,并且存在大体相同的真菌群落. Scagel 等[32]研究表明,臭氧影响北美黄松土壤中活微生物和总微生物的种群组成. 活真菌生物量、 活真菌和细菌生物量比值随植物受臭氧胁迫的时间而增加. 在低臭氧浓度下(23 nL ·L-1),总真菌和总细菌的生物量增加,但臭氧对其的影响不是线性的; 在高臭氧浓度下(31 nL ·L-1),总真菌和总细菌生物量与对照相比是下降的. 由此可见,臭氧改变土壤微生物群落结构的组成,而土壤微生物群落结构中细菌、 真菌和放线菌组成的改变,必然会导致植株营养、 植物竞争和物种组成的改变[18]. 而这种影响将是更加深远且不可逆的,而且对生态系统生产力、 碳循环、 营养循环和水循环将会产生更大更长远的影响.
4 结论(1) 臭氧降低土壤微生物生物量碳,且即时光合产物对地下的输入减少在第一时间即反应出来,第一次标记时虽然总的土壤微生物量碳没有受到臭氧的影响,但臭氧已经降低土壤微生物量中的13 C含量.
(2)臭氧改变了土壤微生物PLFA的结构组成,臭氧胁迫前期,只有细菌脂肪酸的13 C分配受到臭氧的明显影响,而胁迫后期则臭氧主要影响的是13 C对放线菌和真菌的分配.
| [1] | IPCC. IPCC Fourth Assessment Report: Climate Change 2007 (AR4)[EB/OL]. http://www.ipcc.ch/publications_and_data/publications_ipcc_fourth_assessment_report_synthesis_report.htm. |
| [2] | Feng Z W, Jin M H, Zhang F Z, et al. Effects of ground-level ozone (O3) pollution on the yields of rice and winter wheat in the Yangtze River delta[J]. Journal of Environmental Sciences, 2003, 15 (3): 360-362. |
| [3] | Fuhrer J, Booker F. Ecological issues related to ozone: agricultural issues[J]. Environment International, 2003, 29 (2-3): 141-154. |
| [4] | Nunn A J, Kozovits A R, Reiter I M, et al. Comparison of ozone uptake and sensitivity between a phytotron study with young beech and a field experiment with adult beech(Fagus sylvatica)[J]. Environmental Pollution, 2005, 137 (3): 494-506. |
| [5] | Nunn A J, Reiter I M, Hberle K H, et al. Response patterns in adult forest trees to chronic ozone stress: identification of variations and consistencies[J]. Environmental Pollution, 2005, 136 (3): 365-369. |
| [6] | 王雪梅, 符春, 梁桂雄. 城市区域臭氧浓度变化的研究[J]. 环境科学研究, 2001, 14 (5): 1-3, 19. |
| [7] | Ding A J, Wang T, Thouret V. Tropospheric ozone climatology over Beijing: Analysis of aircraft data from the MOZAIC program[J]. Atmospheric Chemistry and Physics, 2008, 8 (1): 1-13. |
| [8] | Wang T, Wei X L, Ding A J, et al. Increasing surface ozone concentrations in the background atmosphere of Southern China, 1994-2007[J]. Atmospheric Chemistry and Physics, 2009, 9 (16): 6217-6227. |
| [9] | Feng Z Z, Zheng H Q, Wang X K, et al. Sensitivity of Metasequoia glyptostroboides to ozone stress[J]. Photosynthetica, 2008, 46 (3): 463-465. |
| [10] | Zhang W W, Niu J F, Wang X K, et al. Effects of ozone exposure on growth and photosynthesis of the seedlings of Liriodendron chinense (Hemsl.) Sarg, a native tree species of subtropical China[J]. Photosynthetica, 2011, 49 (1): 29-36. |
| [11] | Niu J F, Zhang W W, Feng Z Z, et al. Impact of elevated O3 on visible foliar symptom, growth and biomass of Cinnamomum camphora seedlings under different nitrogen loads[J]. Journal of Environmental Monitoring, 2011, 13 (10): 2873-2879. |
| [12] | Skrby L, Ro-poulsen H, Wellburn F A M, et al. Impacts of ozone on forests: a European perspective[J]. New Phytologist, 1998, 139 (1): 109-122. |
| [13] | Chen Z, Wang X K, Feng Z Z, et al. Effects of elevated ozone on growth and yield of field-grown rice in Yangtze River Delta, China[J]. Journal of Environmental Sciences, 2008, 20 (3): 320-325. |
| [14] | Dorhmann A B, Tebbe C C. Effect of elevated tropospheric ozone on the structure of bacterial communities inhabiting the rhizosphere of herbaceous plants native to Germany[J]. Applied and Environmental Microbiology, 2005, 71 (12): 7750-7758. |
| [15] | 寇太记, 于伟伟, 朱建国, 等. 臭氧污染对不同品种小麦干物质与生物量碳积累与分配的影响[J]. 环境科学, 2012, 33 (8): 2862-2867. |
| [16] | Andersen C P, Rygiewicz P T. Allocation of carbon in mycorrhizal Pinus ponderosa seedlings exposed to ozone[J]. New Phytologist, 1995, 131 (4): 471-480. |
| [17] | McCrady J K, Andersen C P. The effect of ozone on below-ground carbon allocation in wheat[J]. Environmental Pollution, 2000, 107 (3): 465-472. |
| [18] | Yoshida L C, Gamon J A, Andersen C P. Differences in above-and below-ground responses to ozone between two populations of a perennial grass[J]. Plant and Soil, 2001, 233 (2): 200-211. |
| [19] | 陈展, 王效科, 段晓男, 等. 臭氧浓度升高对盆栽小麦根系和土壤微生物功能的影响[J]. 生态学报, 2007, 27 (5): 1803-1808. |
| [20] | Chen Z, Wang X K, Feng Z Z, et al. Impact of elevated O3 on soil microbial community function under wheat crop[J]. Water, Air and Soil Pollution, 2009, 198 (1-4): 189-198. |
| [21] | Chen Z, Wang X K, Yao F F, et al. Elevated ozone changed soil microbial community in a rice paddy[J]. Soil Science Society of American Journal, 2010, 74 (3): 829-837. |
| [22] | 王曙光, 侯彦林. 磷脂脂肪酸方法在土壤微生物分析中的应用[J]. 微生物学通报, 2004, 31 (1): 114-117. |
| [23] | 白震, 何红波, 张威, 等. 磷脂脂肪酸技术及其在土壤微生物研究中的应用[J]. 生态学报, 2006, 26 (7): 2387-2394. |
| [24] | 陈晓娟, 吴小红, 刘守龙, 等. 不同耕地利用方式下土壤微生物活性及群落结构特性分析: 基于PLFA和MicroRespTM方法[J]. 环境科学, 2013, 34 (6): 2375-2382. |
| [25] | 王曙光, 冯兆忠, 王效科, 等. 大气臭氧浓度升高对丛枝菌根(AM)及其功能的影响[J]. 环境科学, 2006, 27 (9): 1872-1877. |
| [26] | Lu Y H, Murase J, Watanabe A, et al. Linking microbial community dynamics to rhizosphere carbon flow in a wetland rice soil[J]. FEMS Microbiology Ecology, 2004, 48 (2): 179-186. |
| [27] | US Environmental Protection Agency. Air quality criteria for ozone and related photochemical oxidants. Vo1. Ⅱ. Environmental Protection Agency, Office of Air Quality Planning and Standards, Research Triangle Park, NC. US EPA Report No. EPA 600/P-93/004bF[R]. Washington DC, 1996. |
| [28] | Ariyaphanphitak W, Chidthaisong A, Sarobol E, et al. Effects of elevated ozone concentrations on Thai jasmine rice cultivars(Oryza sativa L.)[J]. Water Air & Soil Pollution, 2005, 167 (1-4): 179-200. |
| [29] | Larson J L, Zak D R, Sinsabaugh R L. Extracellular enzyme activity beneath temperate trees growing under elevated carbon dioxide and ozone[J]. Soil Science Society of America Journal, 2002, 66 (6): 1848-1856. |
| [30] | Shafer S R. Influence of ozone and simulated acidic rain on microorganisms in the rhizosphere of sorghum[J]. Environmental Pollution, 1988, 51 (2): 131-152. |
| [31] | Manning W J, Feder W A, Papia P M, et al. Influence of foliar ozone injury on root development and root surface fungi of pinto bean plants[J]. Environmental Pollution, 1971, 1 (4): 305-312. |
| [32] | Scagel C F, Andersen C P. Seasonal changes in root and soil respiration of ozone-exposed ponderosa pine (Pinus ponderosa) grown in different substrates[J]. New Phytologist, 1997, 136 (4): 627-643. |