 2014, Vol. 35
2014, Vol. 35


污水处理厂尾水细菌群落结构分析
引用本文
徐爱玲, 任杰, 宋志文, 吴等等, 夏岩. 污水处理厂尾水细菌群落结构分析[J]. 环境科学, 2014, 35(9): 3473-3479.
XU Ai-ling, REN Jie, SONG Zhi-wen, WU Deng-deng, XIA Yan. Microbial Community of Municipal Discharges in A Sewage Treatment Plant[J]. Environmental Science, 2014, 35(9): 3473-3479.
污水处理厂尾水细菌群落结构分析
青岛理工大学环境与市政学院, 青岛 266033
摘要:城市尾水中含有大量的微生物,其中相当一部分为病原菌,其排入自然水体后给附近人群造成健康威胁. 本研究采用建立16S rDNA 克隆文库和PCR-RLFP技术对尾水的细菌群落结构进行分析,结果表明尾水中有59种细菌分别属于11个纲,其中44种细菌属于Proteobacteria门,约占85%(其中β-Proteobacteria和γ-Proteobacteria纲占绝对优势),7种细菌属于Clostridia纲占7.2%,而Actinobacteria、Bacteroidia、Gloeobacteria、Nitrospira、Planctomycetacia 这5个纲的细菌所占数量都低于4%,尾水中的细菌大多来源于污水处理过程,因此各地区应该根据受纳水体的特点和净化能力选择合适的污水处理工艺;59种细菌中Legionella spp. 占10%以上,其导致的退伍军人症可能成为尾水排放对附近人群健康的最大威胁,另外尾水中一些细菌能代谢次氯酸盐对氯气有一定抵抗能力,所以在选择消毒措施时应该选用氯气消毒与其他消毒工艺的组合;本研究中粪大肠杆菌、大肠埃希氏菌、沙门氏菌、志贺菌属、霍乱弧菌等致病菌在尾水中均未检出,因此对这些致病菌进行研究需要选择针对性更强的方法.
关键词:
市政尾水
微生物
群落结构
安全
PCR-RFLP
Microbial Community of Municipal Discharges in A Sewage Treatment Plant
School of Environment and Municipal Engineering, Qingdao Technological University, Qingdao 266033, China
Abstract: There are numerous microorganisms, especial pathogens, in the discharges. Those microorganisms are discharged into the river and sea through sewage outfalls, which results in possible health risks to coastal populations. And more attention should be paid to municipal discharges in developing countries. This study investigated the microbial community in the discharges by constructing 16S rDNA clones library and using the PCR-RFLP technology. Phylogenetic analysis of bacteria in municipal discharges showed that there were 59 species, which were divided into 11 classes. Proteobacteria accounted for 85% of all the bacteria, of which β-Proteobacteria and γ- Proteobacteria were the dominant classes. Bacteria in the waste water treating process had important influence on microbial community in municipal discharges, therefore, municipal sewage plant should choose the process according to the characteristics and purifying capacity of the receiving water body. Legionella spp. accounted for approximately 10%, the Legionnaires' disease resulted from which might be of top risk for the residents in the surrounding of the municipal discharges outfall and receiving water. Dechloromonas aromatica could make use of chlorite (ClO-), which led to its survival from chlorine disinfection, and it alerted us that several disinfection methods should be used together to ensure the bacterial safety of municipal discharges. Coliform group and other pathogenic bacteria, such as Salmonella spp., Shigella spp., Escherichia coli, Vibrio cholerae, Staphylococcus aureus, Arcobacter spp. were not detected in this study, and it indicated that we should do more work and use more methods to investigate the perniciousness of discharges.
Key words:
municipal discharges
microbe
community structure
security
PCR-RFLP
污水处理厂对改善水环境状况起到了重大作用,但污水处理厂的尾水出路问题一直未得到有效解决. 许多污水处理厂将尾水直接排入天然水体,巨大的排放量超过天然水体的水环境容量,加剧水环境的污染. 因此尾水问题引起了社会各界的广泛关注,并对尾水深度处理及排放做了大量研究 [1,2],生物滤池、 人工湿地等尾水深度处理技术以及高效尾水排放系统也应运而生. 这些措施在一定程度上降低了尾水排放对自然水体的冲击. 但是在人们关注尾水对自然水体水质产生影响的同时,却忽略了尾水中存在的大量微生物对自然水体生态平衡及附近人群可能造成的危害.
城市污水中含有大量的微生物,其中相当一部分为病原菌,如: 沙门氏菌(Salmonella spp.)、 志贺氏菌(Shigella spp.)、 大肠埃希氏菌(Escherichia coli)、 霍乱弧菌(Vibrio cholerae)、 金黄色葡萄球菌(Staphylococcus aureus)、 弓形杆菌(Arcobacter spp.)等[3,4],可对人身体健康和社会经济造成严重危害. 研究发现这些病原菌主要是人源污水微生物,污水处理工艺对其有一定去除效果[5],消毒可使病原菌的数量大大降低[6],但有些微生物生存能力强,能够产生芽孢和孢子,难以彻底去除. 目前对城市污水的微生物学评价多采用细菌总数和总大肠杆菌数,但粪大肠菌群与其他尾水常见病原菌相关性较低,而不能有效代表全部的微生物或病原菌[7]. 国内对尾水安全的研究起步较晚,主要集中于消毒、 尾水排放对受纳水体影响和部分回用水的微生物安全研究[8, 9, 10]. 文献[11,12]采用定量聚合酶链式反应技术(qPCR)建立了尾水中某些病原菌的定量检测方法,证实了尾水中病原菌定量检测的可行性. 但均未对尾水微生物的群落结构进行详细的分析.
传统的微生物培养方法难以获得尾水微生物群落结构的准确信息[13],随着现代分子生物学的发展,出现了一些更快、 更准确分析微生物群落结构及多样性的方法,这些方法突破了传统研究方法要依赖于培养技术的局限而显示出明显的优越性. 研究证实RFLP技术具有较高的分辨率和可重复性,已广泛用于环境中微生物的群落结构分析[14, 15, 16, 17, 18, 19]. 因此,本研究拟采用PCR-RLFP技术对青岛市某污水处理厂待排尾水的微生物群落结构进行分析,以期为尾水排放安全防范措施的提出提供依据,对于公共健康有着十分重要的意义. 1 材料与方法 1.1 主要试剂和仪器
试剂:土壤DNA提取试剂盒(OMEGA),DNA凝胶回收试剂盒(OMEGA),PCR mix,内切酶,琼脂糖,TAE缓冲液(50×). 仪器:PCR仪,凝胶成像系统(gene公司); 高速冷冻离心机,超净工作台,核酸超微量分析仪. 1.2 样品采集
2013-06-10采用击开式采水器采集青岛市某污水处理厂待排尾水水样,低温保存并在2 h内送至实验室进行抽滤,抽滤后细菌用于基因组提取. 该污水处理厂以A2O工艺为主要处理工艺,采用次氯酸钠消毒,日处理水量1.7×105 m3,其中60%为工业废水,40%为生活污水. 进出水水质指标如表 1.
| 表 1 污水厂进出水水质指标 Table 1 Water quality indexes of waste water and treated waste water |
采用土壤DNA提取试剂盒(OMEGA)的方法,获得纯度和含量较高的DNA模板,可进一步供PCR扩增使用. DNA检测使用1%琼脂糖进行水平电泳,在0.5 g ·mL-1的溴化乙锭溶液中染色30~45 min,用去离子水漂洗后紫外下观察. 1.4 16S rDNA基因的PCR扩增
两个引物用于扩增16S rRNA基因片断,上游引物27F (5′-AGAGTTTGATCCTGGCTCAG-3′) 和下游引物1492R(5′-GGTTACCTTGTTACGACTT-3′)[20]. 通过低熔点琼脂糖凝胶电泳观察PCR结果,使用Wizard DNA Clean-Up Kit (Promega)试剂盒回收目的片断. 1.5 16S rDNA文库的构建与筛选
将纯化后的DNA片段通过T4连接酶克隆试剂盒(MBI Fermentas)连接在Ptz57R/T载体上,转化到感受态大肠杆菌(DH5α)细胞中,涂布于加有50 μg ·mL-1 Amp及IPTG、 X-gal的生色LB琼脂平板上,放在37℃培养箱中正培养2 h,再倒置培养10~16 h,培养基平板上长出直径3~5 mm的菌落. 随机挑选平板中的200个阳性克隆子,采用菌体直接扩增方式,用pTZ57R/T Vector通用引物M13/PUCR 5′-3′:GTAAAAC GACGGCCAGT; M13/PUCF 5′-3′:CAGGAAACAGCTATGAC扩增外源插入片段,重新获得目标基因. 1.6 阳性克隆子的PCR-RFLP分析
扩增产物用4碱基限制性内切酶HhaI消化从各克隆子扩增出来的目标片段. 用3%琼脂糖凝胶分离酶切片段,200 V下电泳2.5 h,用凝胶成像仪拍照,并对凝胶电泳图谱进行分析,将每个酶切分型结果作为一个OTU(operational taxonomic unit). 1.7 测序和构建系统发育树
选取PCR-RFLP筛选出的阳性克隆子,送上海生物工程公司进行序列测定. 测序所得16S rRNA序列,采用Blast软件在GenBank进行相似性搜索(http://www. ncbi. nlm. nih. gov. blast),获取近似序列及对应菌种信息. 用Clustal X程序进行比对,用邻接法(Neighbor-joining Method)构建系统发育树. 拓扑分析为1000次重复取样结果,生成发育树采用Treeview重建. 1.8 分析方法
(1)文库的库容(coverage)计算公式为:
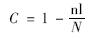
式中,N代表 16S rDNA文库总克隆数,nl代表在文库中仅出现一次的OTUs(operational taxonomic units)的数量.
(2)Shannon-Wiene多样性指数(H′):
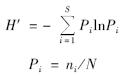
式中,S为16S rDNA的所有RFLP总类型数,ni为第i种16S rDNA的RFLP变异类型克隆数,N为总克隆数.
(3)丰富度指数 (Margalef):
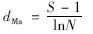
(4)均匀度指数的计算公式为:
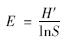 2 结果与分析
2.1 基因提取及16S rDNA扩增
2 结果与分析
2.1 基因提取及16S rDNA扩增
尾水化学组分复杂,能否成功提取目标DNA片段及提取效率的高低直接影响后续RFLP分析. 从样品中提取的DNA通过琼脂糖凝胶电泳检验,得到一条清晰条带,与细菌基因组的大小相同,因此所提DNA属于比较完整的细菌基因组DNA. 且条带的亮度和清晰度较好没有出现拖尾现象,说明DNA的纯度较高,提取获得了满意的结果[21]. 经PCR反应,获得了样品中细菌16S rDNA的目的片段,长约1500 bp且亮度和纯度都比较好,未出现非特异性扩增,且阴性对照未有产物出现,说明PCR扩增效果良好. 2.2 16S rDNA基因的PCR扩增
图 1为克隆子扩增的琼脂糖凝胶电泳图谱,从中可以看出,条带长约1600 bp,亮度和纯度都比较好,克隆率较高且未出现非特异性扩增,可以用于后续实验.
 | 1~61:可用于酶切的克隆子16S rDNA扩增片段 图 1 部分克隆子16S rDNA扩增的琼脂糖凝胶电泳图谱 Fig. 1 Agarose gel electrophoresis of part of the 16S rDNA fragments amplified from clones |
从文库中共挑取白色菌落200个,其中阳性克隆168个,阳性率83.3%. 阳性克隆子 M13 PUC引物的PCR产物HhaⅠ酶切图谱如图 2所示,用Quantity one凝胶分析软件对RFLP图谱分析表明,168个阳性克隆中存在59个不同类型的OTUs. 其中A33的OTU占7.14%,比例超过3%的OTUs一共有9种,仅有唯一克隆子的OUTs有18个. 该文库的生物多样性指数如表 2所示. 克隆文库的覆盖度(coverage value,C)体现样品中微生物的覆盖程度,其数值越大,所构建的克隆文库包含的样品中的微生物种类就越多,因此越能真实地反映该样品中微生物多样性特征. 但是Kemp等[22]研究发现即使再大的克隆文库都不能囊括环境样品中的所有微生物,理论上来讲,当覆盖度超过80%即可认为该克隆文库具有较高的库容. 本研究中尾水克隆文库库容值C达到89.3%,表明所研究的克隆文库库容比较高,能真实地反映样品中的细菌群落结构组成. 丰富度指数(dMa)是与物种均匀度无关的多样性指数,它仅与环境中物种数目的多寡有关,本研究中dMa为 11.3,说明本样品中物种数量较多. 香农多样性指数(Shannon Diversity Index,H′)在均一度存在明显差异的群体中存在一定的缺陷,即低丰富度和高均匀度的群落与具高丰富度和低均匀度的群落多样性指数可能相同[23],但对某一指定样品却是能反映丰富度和均匀度的综合指标,本克隆文库中H′为3.84,说明尾水中细菌丰富度和均匀度较高. 均匀度(species evenness,E)是指生境中全部物种个体数目的分配状况,它反映的是各物种个体数目分配的均匀程度,本研究中尾水的均匀度指数为75%,说明尾水中细菌分布相对均匀,优势物种种类和数量不多.
 | 图 2 部分克隆子 M13 PUC引物的PCR产物HhaⅠ酶切图谱 Fig. 2 Agarose gel electrophoresis of part of the restriction fragment length profiles of 16S rDNA fragments amplified from discharges |
| 表 2 尾水克隆文库多样性指数 Table 2 Diversity index of the clone library from wastewater |
2.4 克隆子测序及系统发育学分析
对经过RFLP分析获得的59个克隆的插入序列进行序列测定. 一般来说,当微生物的16S rDNA同源性达到97%或以上时,可以将这些菌划为一个种; 同源性达到94%或以上时,可以将这些菌划为一个属[24]. 因此采用BLAST软件对所获得克隆子序列与迄今已描述的原核生物16S rDNA 序列相似性进行了比对(表 3). 59个克隆子序列中仅有30个与已知物种相似度在94%以上,10种相似度低于90%,说明尾水中仍有许多未经纯培养得到分离鉴定的物种,因此对尾水的生物安全性认识也较为不足. 59个物种中,Legionella longbeachae占7.14%,具有一定优势,Pseudomonas kilonensis、Dechloromonas aromatica、Desulfococcus multivorans 所占比例均大于4%,也属于相对优势物种.5个物种所占比例介于4% ~3%之间,6个物种所占比例介于3% ~2%,26个物种所占比例在2% ~1%之间,18个物种所占种比例均低于1%.
| 表 3 样品中细菌16Sr DNA的BLAST分析结果 Table 3 Inventory of bacterial 16S rDNA fragments classified into groups according to ARDRA patterns and sequence similarity |
59种细菌分别属于11个纲(图 3). 44种细菌属于变形菌门(Proteobacteria)约占85%,其中23种属于β-变形菌纲占41.4%,10种属于γ-变形菌纲占26.5%,α-变形菌纲有5种细菌,δ-变形菌纲有3种细菌,这一结果与尾水排放口附近的微生物群落结构组成极为相似[25],说明尾水对受纳水体生物多样性具有重要的影响. 7种细菌属于Clostridia纲占7.2%,而Actinobacteria、 Bacteroidia、 Gloeobacteria、 Nitrospira、 Planctomycetacia 这5个纲的细菌所占数量都低于4%.
 | 图 3 尾水中不同纲细菌所占比例
Fig. 3 Classification and composition of species in the discharge
|
Legionella longbeachae最早是在1981年时从肺炎患者体内分离到的一类革兰氏阴性细菌,可引起肺炎[23]. 虽然已报道的几例肺炎患者,都是直接与被Legionella longbeachae污染的土壤或者肥料接触引起的,但是新的传播载体有可能导致该传染病的暴发[26]. 沿海城市尾水一般都会直接排入附近海域,虽然高盐环境和光照具有一定的杀菌作用[7],但是新的环境也有可能存在Legionella longbeachae的新传播载体. 尾水中除了Legionella longbeachae,还存在Legionella rubrilucens、 Legionella impletisoli,致使Legionella spp.所占比例超过10%,由于污水处理厂巨大的排放量,Legionella spp.引发的退伍军人症将会对附近人群健康产生潜在的巨大威胁.
尾水中Pseudomonas kilonensis、 Dechloromonas aromatica、 Desulfococcus multivorans、 Azospira restricta、 Denitratisoma oestradiolicum、 Massilia aurea、 Ralstonia eutropha、 Tistrella mobilis所占比例均超过3%,这8类细菌均在水处理系统中起着重要的作用并占一定的比例[27],例如:Azospira restricta 具有氧化催化能力,能积累PHA,兼具脱氮除磷的作用,是一种典型的反消化除磷菌; Hydrogenophaga defluvi有厌氧消化能力,在水处理脱氮过程中有重要的作用,它还能利用有机酸、 氨基酸和蛋白胨等含氮化合物,但是对糖类的利用能力较低; Dechloromonas aromatic 可以代谢苯,同时利用碳源作为电子受体,因此在脱氮除磷中起着非常重要的作用. 由此可见水处理微生物对尾水微生物群落结构有重要的影响,而水处理微生物的种类除受进水水质影响外,主要取决于污水处理工艺,不同的污水处理工艺会导致水处理系统微生物群落结构的差异,因此各城市污水处理厂应该根据受纳水体的特点和净化能力选择适合的污水处理工艺.
为降低尾水中细菌尤其是致病菌的数量,在其排入自然水体前通常要进行消毒后处理,常用的消毒方式主要有紫外、 氯气和臭氧,消毒后尾水中微生物的数量大大降低(99.9%),但有些微生物生存能力强,难以彻底去除[7]. 本研究发现尾水中含有4%以上的 Dechloromonas aromatic,该菌有次氯酸盐歧化酶和血红素加氧酶,前者能迅速地将次氯酸盐(ClO-)分解成氯离子(Cl-)和氧气,后者能以次氯酸盐(ClO-)作为氧化剂参加氧化还原反应[24],因此它具有一定的代谢次氯酸盐的能力,这可能是Dechloromonas aromatic在尾水中有较大比例的一个重要原因. 这也提示污水处理厂在选择消毒工艺时使用不同工艺的组合可大大提高尾水中微生物的除去效果. 粪大肠杆菌、 大肠埃希氏菌、 沙门氏菌、 志贺菌属、 霍乱弧菌等致病菌均未检出,这主要是因为这些细菌数量较少所占比例较低,而本研究所建的克隆文库不够大所致,因此有对这些致病菌进行研究需要选择更加针对性的研究方法进行检测.
为保护重点湖泊、 水库、 河段,世界不少沿海国家和地区常常将尾水调离敏感水域,集中输送至海洋进行最终处置[28]. 目前尾水排海主要有深海排放和近岸直排两种方式,深海排放对环境影响较小,但深海排污工程造价高,一般为发达国家或者发展中国家的特定地区所采用. 目前我国大部分沿海地区一般在尾水达到一级A或者B标准后采用近岸直排的方式. 近岸水深较浅,流速较慢,再加上排海尾水量日益增加,致使微生物尤其是病原微生物难以得到较快的输送和消灭而被聚集在近岸海域[22],本研究发现尾水中存在的大量Legionella spp.,其引起的疾病可能会对排水口附近海水浴场人群的健康造成威胁. 同时尾水中含有超过50种微生物,大量异源微生物入侵海洋也会影响海洋生态环境,造成海水养殖损失,因此尾水的处置不容忽视.
4 结论
(1)尾水中含有59种细菌分别属于11个纲,其中 44种细菌属于变形菌门(Proteobacteria)约占85%(β-变形菌纲和γ-变形菌纲占绝对优势),7种细菌属于Clostridia纲占7.2%,而Actinobacteria、 Bacteroidia、 Gloeobacteria、 Nitrospira、 Planctomycetacia 这5个纲的细菌所占数量都低于4%. 这些微生物大多来源于污水处理过程,因此各地区应该根据受纳水体的特点和净化能力选择合适的污水处理工艺.
(2) 尾水的59种细菌中Legionella spp.占10%以上,其导致的退伍军人症可能成为尾水排放对附近人群健康的最大威胁; 另外一些细菌能代谢次氯酸盐对氯气有一定抵抗能力,所以在选择消毒措施时应该选用氯气消毒与其他消毒工艺的组合. 本研究中粪大肠杆菌、 大肠埃希氏菌、 沙门氏菌、 志贺菌属、 霍乱弧菌等致病菌在尾水中均未检出,因此对这些致病菌进行研究需要选择针对性更强的方法.
参考文献
| [1] | 杨林, 李咏梅. 组合人工湿地处理工业园区污水厂尾水的中试研究[J]. 环 境 工 程 学 报, 2012, 6 (6): 1846-1850. |
| [2] | 李魁晓, 白雪, 李鑫玮, 等. 城市污水厂二级处理出水深度处理组合工艺研究[J]. 环境工程学报, 2012, 6 (1): 63-67. |
| [3] | Ho H T, Lipman L J, Gaastra W. Arcobacter, what is known and unknown about a potential foodborne zoonotic agent[J]. Veterinary Microbiology, 2006, 115 (l-3): l-13. |
| [4] | He X H, Ma S H, Li A D. Concentration limitations of pathogenic microorganisms in reclaimed water [J]. Environmental Science, 2006, 27 (7): 1402-1405. |
| [5] | Cai L, Ju F, Zhang T. Tracking human sewage microbiome in a municipal wastewater treatment plant[J]. Applied Microbiology and Biotechnology, 2014, 98 (7): 3317-3326. |
| [6] | Eddabra R, Moussaoui W, Prévost G,, et al. Occurrence of Vibrio cholerae non-O1 in three wastewater treatment plants in Agadir (Morocco) [J]. World Journal of Microbiology and Biotechnology, 2011, 27 (5): 1099-1108. |
| [7] | Wéry N, Lhoutellier C, Ducray F, et al. Behaviour of pathogenic and indicator bacteria during urban wastewater treatment and sludge composting, as revealed by quantitative PCR[J]. Water Research, 2008, 42 (1-2): 53-62. |
| [8] | 王玲玲. 城市再生水用于循环冷却系统水中微生物腐蚀控制[D]. 天津: 天津大学, 2011. |
| [9] | 郭逍宇, 董志, 宫辉力. 再生水灌溉对草坪土壤微生物群落的影响[J]. 中国环境科学, 2006, 26 (4): 482-485. |
| [10] | 谢兴, 胡洪营, 郭美婷, 等. 再生水雾化导致的病原微生物暴露剂量计算方法研究[J]. 环境科学, 2009, 30 (1): 70-74. |
| [11] | 刘永军, 张崇淼, 王晓昌, 等. 环境水体中肠道病原细菌的定量PCR检测[J]. 环境科学, 2008, 29 (5): 1175-1180. |
| [12] | 张崇淼, 王晓昌, 周进宏, 等. 城市地表水中肠道病原微生物与粪便污染指示菌的关系研究[J]. 环境科学学报, 2012, 32 (11): 2789-2794. |
| [13] | Eschenhagen M, Schuppler M, Rske I. Molecular characterization of the microbial community structure in two activated sludge systems for the advanced treatment of domestic effluents[J]. Water Research, 2003, 37 (13): 3224-3232. |
| [14] | Regan J M, Harrington G W, Noguera D R. Ammonia-and nitrite-oxidizing bacterial communities in a pilot-scale chloraminated drinking water distribution system [J]. Applied and Environmental Microbiology, 2002, 68 (1): 73-81. |
| [15] | Mintie A T, Heichen R S, Cromack K Jr, et al. Ammonia-oxidizing bacteria along meadow-to-forest transects in tne oregon cascade mountains[J]. Applied and Environmental Microbiology, 2003, 69 (6): 3129-3136. |
| [16] | Park H D, Noguera D R. Evaluating the effect of dissolved oxygen on ammonia-oxidizing bacterial communities in activated sludge[J]. Water Research, 2004, 38 (14-15): 3275-3286. |
| [17] | Bernhard A E, Donn T, Giblin A E, et al. Loss of diversity of ammonia-oxidizing bacteria correlates with increasing salinity in an estuary system[J]. Environmental Microbiology, 2005, 7 (9): 1289-1297. |
| [18] | Yeager C M, Northup D E, Grow C C, et al. Changes in nitrogen-fining and ammonia-oxidizing bacterial communities in soil of a mixed conifer forest after wildfire[J]. Applied and Environmental Microbiology, 2005, 71 (5): 2713-2722. |
| [19] | Cabezas A, Draper P, Muxí L, et al. Post-treatment of a slaughterhouse wastewater: stability of the microbial community of a sequencing batch reactor operated under oxygen limited conditions [J]. Water Science and Technology, 2006, 54 (2): 215-221. |
| [20] | Xu A L, Xia J L, Zhang S, et al. Bioleaching of chalcopyrite by UV-induced mutagenized Acidiphilium cryptum and Acidithiobacillus ferrooxidans [J]. Transactions of Nonferrous Metals Society of China, 2010, 20 (2): 315-321. |
| [21] | 李鹏. 城市污水生物脱氮除磷活性污泥系统微生物多样性研究[D]. 青岛: 中国海洋大学, 2007. |
| [22] | Kemp P F, Aller J Y. Bacterial diversity in aquatic and other envirunments: What 16S rDNA libraries Cal tell US[J]. FEMS Microbiology Ecology, 2004, 47 (2): 161-177. |
| [23] | 白玉涛, 玉周, 吉赵. 内蒙古高原干涸湖泊反硝化及甲烷氧化细菌的群落分析[J]. 中国环境科学, 2012, 32 (7): 1293-1301. |
| [24] | Yang L, Chang W S, Huang M N L. Natural disinfection of wastewater in marine outfall fields[J]. Water Research, 2000, 34 (3): 743-750. |
| [25] | Yang Y, Wang X, Shi J F, et al. The influence of the discharging sewage on microbial diversity in sludge from Dongting Lake[J]. World Journal of Microbiology Biotechnology, 2012, 28 (2): 421-430. |
| [26] | 孙寓姣, 左剑恶, 陈莉莉. 同时产甲烷反硝化颗粒污泥中微生物群落结构[J]. 中国环境科学, 2007, 27 (1): 44-48. |
| [27] | 韩甜甜. 复合铁酶促活性污泥工艺生物脱氮除磷效能及微生物多样性研究[D]. 青岛: 青岛理工大学, 2011. |
| [28] | Aguilar M I, Sáez J, Lloréns M, et al. Nutrient removal and sludge production in the coagulation-flocculation process[J]. Water Research, 2002, 36 (11): 2910-2919. |