 2014, Vol. 35
2014, Vol. 35



2. 同济大学污染控制与资源化研究国家重点实验室,上海 200092;
3. 西南科技大学固体废物处理与资源化教育部重点实验室,绵阳 621010
2. State Key Laboratory of Pollution Control and Resource Reuse, Tongji University, Shanghai 200092, China;
3. Key Laboratory of Solid Waste Treatment and Resource Recycle, Ministry of Education, Southwest University of Science and Technology, Mianyang 621010, China
铬(Cr)自1797年由法国科学家Vauquelin从铬铅矿中发现以来[1],被广泛地应用于皮革、 电镀、 颜料、 木材防腐等行业,为工业发展做出了巨大贡献[2].但与此同时,六价的氧化形式Cr(Ⅵ)由于具有强烈的生物毒性和极强的水相迁移性[3],造成了日益严重的环境污染.而Cr(Ⅲ)在环境中易形成非晶型的氢氧化物沉淀,且生物毒性较小(由于细胞膜对游离Cr(Ⅲ)的不可渗透性,使得其毒性仅为Cr(Ⅵ)的千分之一)[4],因此将Cr(Ⅵ)还原成Cr(Ⅲ)已成为一种公认的治理Cr(Ⅵ)污染的有效策略.自从20世纪70年代Romanenko等[5]首次在厌氧环境中发现了具有六价铬还原能力的细菌以来,细菌介导的Cr(Ⅵ)还原由于具有反应温和、 能耗低且无二次污染的特点,被认为是最具潜力的Cr(Ⅵ)环境修复方法之一[6, 7].尽管大量具有Cr(Ⅵ)还原功能的细菌被筛选出来,一些参与还原过程的功能蛋白及其编码基因也被研究,但是Cr(Ⅵ)细菌还原的机制尚不清晰,比如还原过程中的电子转移、 蛋白表达及调控等,这些都亟需深入研究.因此,探寻细菌Cr(Ⅵ)还原的机制,尤其是从分子水平上了解还原过程中分子机制,就有可能抓住制约其还原效率的关键环节,以期为Cr(Ⅵ)污染的细菌修复提供科学依据. 1 Cr(Ⅵ)还原细菌
目前已报道的具有Cr(Ⅵ)还原功能的细菌类群主要分布(见表1)在:①变形菌门(Proteobacteria)下的α-、 β-、 γ-和δ-变形菌纲; ②厚壁菌门(Firmicutes)下的芽孢杆菌纲和梭菌纲; ③放线菌门(Actinobacteria)下的放线菌纲; ④栖热菌门(Deinococcus-Thermus)下的异常球菌纲.其中广泛用于分子机制研究的模式菌株有:Shewanella oneidensis MR-1、 P. putida PRS 2000、 P. putida MK1、 P. ambigua G-1、 E. coli ATCC 33456、 B. megaterium TKW3、 T. scotoductus SA-01和Desulfovibrio vulgaris Hidenborough等,它们是已被证实携带具有Cr(Ⅵ)还原基因的样本菌株,通过研究这些模式菌株及其在反应过程中表现出来的各种现象,Cr(Ⅵ)还原的大致途径被推测为直接还原与间接还原两种.
| 表1 Cr(Ⅵ)还原细菌及其特性 Table 1 Cr(Ⅵ)-reducing bacteria and their characteristics |
Cr(Ⅵ)直接还原是指细菌通过蛋白的各种作用(如酶的催化作用,功能蛋白的电子传递作用等)将Cr(Ⅵ)直接还原成Cr(Ⅲ),从而降低对细胞的毒性.根据是否有氧气的参与,直接还原可以分为好氧直接还原和厌氧直接还原两种类型.
(1)好氧直接还原在氧气存在的情况下Cr(Ⅵ)还原成Cr(Ⅲ)通常有2步或3步,伴随着短暂中间产物Cr(Ⅴ)和Cr(Ⅳ)的生成.研究表明,NAD(P)H及细胞内存储的电子经ChrR作用,由单电子穿梭形成瞬态的Cr(Ⅴ),接着再经历2个电子传递生成Cr(Ⅲ),虽然部分中间体Cr(Ⅴ)的再氧化会形成活性氧自由基(reactive oxygen species,ROS)对细胞产生一定的毒性,但紧接其后的2个电子传递降低了产生自由基的几率[66, 67].YieF催化Cr(Ⅵ)直接还原成Cr(Ⅲ),这个过程涉及4个电子的传递,其中3个在Cr(Ⅵ)还原中被消耗,另外1个传递给了氧.YieF产生的ROS量最少,因此被认为其Cr(Ⅵ)还原作用比ChrR更加有效[68].以研究较多并具有代表性的ChrR和YieF为例,提出了Cr(Ⅵ)细菌好氧直接还原途径[图1(a)].由电化学原理可知,Cr(Ⅵ)的电极电势比O2更高,因此,在好氧条件下,Cr(Ⅵ)将会优先作为电子受体被还原成Cr(Ⅲ),而Cr(Ⅲ)在环境中通常是以Cr(OH)3的形式存在,且很难被O2重新氧化为Cr(Ⅵ).文献表明,有H2 O和O2存在,温度300℃且反应超过12 h的条件下,Cr(Ⅲ)也只有50%转化为Cr(Ⅵ)[69],由此可见,自然环境下,O2不会抑制细菌的Cr(Ⅵ)还原作用.
 | 图1 Cr(Ⅵ)细菌 Fig.1 Bacterial direct reduction of Cr(Ⅵ) |
(2)厌氧直接还原厌氧环境下的Cr(Ⅵ)还原与呼吸链电子传递相关,以碳水化合物、 蛋白、 脂肪、 氢气和NAD(P)H等作为电子供体,Cr(Ⅵ)为最终电子受体,通过细胞色素家族(如细胞色素b和细胞色素c)进行电子传递完成Cr(Ⅵ)还原.在此过程中,可溶性的以及膜相关的蛋白都被发现参与Cr(Ⅵ)的还原,根据电势的高低,电子的传递途径应该是:NADH脱氢酶 泛醌(UQ) 细胞色素bc1复合体 细胞色素c 细胞色素aa3氧化酶[70].Desulfovibrio vulgaris Hidenborough的全基因组序列分析表明[71],细胞周质中氢气氧化产生的电子有可能暂时储存在multiheme细胞色素c3里,直到这些电子通过电子穿梭体透过内膜.Tetrahemic细胞色素c3(DVU3171)被认为是最初的电子受体,并且其构成了周质中细胞色素c的主要部分.不同的细胞色素c之间的电子传递构建了一个巨大的相互关联的血红素网络.这个网络可能提供了连接多种周质氧化还原蛋白的“电线”,同时还可作为储存来自于氢或甲酸氧化的低电势电子的“电容”.更为重要的是,这个网络提供了Cr(Ⅵ)还原所需要的电子[图1(b)]. 2.2 间接还原
在细菌的厌氧代谢过程中,以硫酸盐还原菌(SRB)为代表的细菌不仅可以通过上述厌氧途径直接还原,还可以通过硫酸盐代谢,生成S2-,S2-再与Cr(Ⅵ)发生氧化还原反应,从而实现Cr(Ⅵ)的间接还原,反应方程式[72]如下所示:
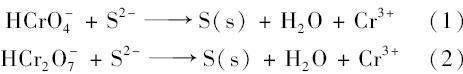
还有报道称氧化亚铁硫杆菌(Acidithiobacillus ferrooxidans)能够通过硫的代谢生成还原态的硫,还原态的硫对Cr(Ⅵ)进行间接还原[20],但是,由于该反应体系要用还原态的硫作为反应基质,因此无法区分到底是细菌的间接作用,还是Cr(Ⅵ)直接氧化了还原态硫基质,该反应机制还有待进一步的验证. 2.3 还原过程中的电子供体及碳源
直接还原中细菌不仅可利用乙酸、 乳酸、 苹果酸、 琥珀酸、 乙醇、 甘油、 葡萄糖等小分子碳源作为电子供体,还可以H2、 NAD(P)H、 AQDS、 偶氮燃料(Azo)作为电子供体8,9,10,11,12,1314,15,16,17,18,19, 21,22,23,24,25,26,27,28,29,30,31,32, 36,37,3839,40,41,42,43,44,45,46,47,48, 49,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65].而间接还原首先是SO2-4作为电子受体,甲酸盐、
乙酸盐、 丙酸盐、 乳酸盐、 丙酮酸盐、 苹果酸盐、 葡萄糖、 乙醇、 乙酸、 丁酸等既是碳源也是电子供体[33,34,35, 50],另外,H2是硫酸盐还原过程中一个重要的电子供体,当该过程生成的H2S与Cr(Ⅵ)反应时,S2-作为电子供体,Cr(Ⅵ)作为最终的电子受体.因此,很多碳源在Cr(Ⅵ)细菌还原过程中充当着电子供体的角色,合理地控制碳源的种类和比例是提高Cr(Ⅵ)细菌还原效率的一种重要途径,在工程实际应用中具有重要的意义. 3 Cr(Ⅵ)还原相关基因研究
专门针对Cr(Ⅵ)还原相关基因簇的研究
未见报道,因为这些基因的主要功能不是进行Cr(Ⅵ)还原,而是参与细菌呼吸链上的电子传递,具有Cr(Ⅵ)还原功能的外膜多铁血红素细胞色素c蛋白MtrC,其编码基因mtrC就位于Shewanella oneidensis MR-1中编码电子传递功能的基因簇上,该基因簇上的mtrA和mtrB编码的MtrA和MtrB,与MtrC共同组成了电子传递的复合蛋白,使得电子可以在细胞内外进行传递[73],而另一种具有Cr(Ⅵ)还原功能的细胞色素c类的蛋白OmcA的编码基因omcA也位于该基因簇上,如图2所示.
 | 图2 Shewanella oneidensis MR-1的Cr(Ⅵ)还原相关的细胞色素c基因簇示意
Fig.2 A schematic diagram of c-type cytochrome gene cluster about Cr(Ⅵ) reduction in Shewanella oneidensis MR-1
|
编码黄素蛋白类铬酸盐还原酶ChrR的基因chrR研究较多,在Escherichia coli str. K-12 substr. MG1655中该基因之前被称为yieF,其编码的蛋白能直接还原Cr(Ⅵ).NCBI的数据库中的信息显示Escherichia coli其它菌株(str. K-12 substr. W3110,BW2952,O127:H6 str. E2348/69,str. K-12 substr.DH10B等)由yieF编码的蛋白具有Cr(Ⅵ)还原功能.关于黄素蛋白类Cr(Ⅵ)还原的基因簇及表达调控机制的研究还鲜见报道,主要是因为其不受铬酸盐诱导,这些酶的活性可能是偶然的 4 Cr(Ⅵ)还原相关蛋白
从Cr(Ⅵ)细菌还原途径可以看出,蛋白在还原过程中起着关键的作用,尤其是能直接催化还原Cr(Ⅵ)的蛋白显得颇为重要,目前已知的具有Cr(Ⅵ)还原功能的蛋白主要有以下几种类型(表2).
| 表 2 具有Cr(Ⅵ)还原能力的蛋白及特性1)Table 2 Cr(Ⅵ)reducing proteins and their characteristics |

从表2可以看出研究得比较多的有细胞色素c类和黄素蛋白类,此外还有老黄酶类等.在这几类蛋白中研究得比较深入,且晶体结构得到详细解析的有:细胞色素c类的Cyt c7,黄素蛋白类的ChrR及老黄酶类的CrS.这3种蛋白在细菌直接还原Cr(Ⅵ)的分子机制中非常具有代表性,广泛地参与了厌氧和好氧两种途径的还原过程,因此其特征和功能的解析对深入揭示Cr(Ⅵ)细菌还原的分子机制研究非常重要. 4.1 Cyt c7
硫和硫酸盐还原细菌不仅可以产生H2S等可以间接还原重金属离子的物质,还能通过酶催化还原Cr(Ⅵ)等金属离子.目前发现在这类细菌中Cr(Ⅵ)是由多血红素细胞色素蛋白催化还原的,例如已发现的四血红素细胞色素c3(Cyt c3)和三血红素细胞色素c7(Cyt c7)均属于细胞色素c3超家族[77]. Desulfomicrobium norvegicum和Desufovibrio vulgaris strain Hildenborough中含有四血红素Cyt c3,可以还原Cr(Ⅵ).核磁共振结果表明Desulfuromonas acetoxidans中的Cyt c7由68个氨基酸残基组成并含有3个顺式亚铁血红素,其蛋白折叠与四血红素Cyt c3很类似.通过研究从Desulfuromonas acetoxidans中获得的Cyt c7,采用核磁共振(NMR)的方法, 初步找到了其还原CrO2-4的机制,Cyt c7蛋白结构中含有hemes Ⅰ、 hemes Ⅱ和hemes Ⅳ这3个部分,其中hemes Ⅳ能给铬提供电子,因为hemes Ⅳ距离CrO2-4在Cyt c7上绑定的位置非常近,通过解读NMR谱,研究者发现整个Cr(Ⅵ)还原过程主要分3步:①铬酸盐和完全还原的细胞色素c形成复合物,铬酸盐阴离子通过静电反应和距离heme Ⅳ很近的赖氨酸形成的正电区域结合; ②结合之后通过3电子传递,将Cr(Ⅵ)还原成Cr(Ⅲ),形成的Cr(Ⅲ)再和赖氨酸进行绑定; ③当部分Cyt c7氧化时,这些蛋白分子之间会进行电子的重新分配,Cyt c7的位点将重新具有对Cr(Ⅵ)的电子传递能力,Cyt c7的结构如图3所示[75, 76].
4.2 ChrR 目前已经在Escherichia coli、 Pseudomonas putida和Gluconacetobacter hansenii中均发现了具有铬酸盐还原能力的ChrR[84].该蛋白属于可溶性黄素蛋白家族,与黄素单核苷酸(FMN)牢固结合,以四聚体形式存在,以NAD(P)H作为电子供体行使双电子还原酶功能.催化Cr(Ⅵ)还原为Cr(Ⅲ)过程中会产生Cr(Ⅳ)和Cr(Ⅴ)中间体. Eswaramoorthy等[79]于2012年解析了Escherichia coli BL21(DE3)中ChrR的晶体结构,该蛋白的晶体为四聚体[图4(c)],四聚体形式具有铬酸盐还原作用.在每个单体C端折叠的1、 3和4环组成的浅凹处,均有1个FMN以多个氢键与蛋白结合[图4(a)和(b)],并且该处含有1个NAD(P)H黄素单核苷酸还原酶的特征氨基酸序列([图4(b)]中带下划线的为氨基酸残基). 2个二聚体由两对氢键组成四聚体,其中1个二聚体的Tyr128和Glu146与另外1个二聚体的Arg125和Tyr85形成氢键,以上4个氨基酸残基的任何1个的变化均会提高该酶的六价铬还原能力,说明这2对氢键组成的网络结构可能属于铬酸盐还原的催化中心. 4.3 CrS

Opperman等[85]于2008年从南非金矿中分离到的Thermus scotoductus SA-01菌株可以还原包括六价铬在内的多种金属.该菌中的铬酸盐还原酶(CrS)是一类新的具有热稳定性的铬酸盐还原酶,属于OYE(old yellow enzyme)家族,单体相对分子质量约36000,含有1个非共价结合的黄素单核苷酸辅因子.该酶最适pH为6.3,最适温度65℃,催化活性需要Ca2+和Mg2+离子,以NAD(P)H作为电子供体.在好氧和厌氧条件下,还原1 mol六价铬分别需要2 mol和1.5 mol的NAD(P)H. 2010年Opperman等[81]解析了CrS全酶的晶体结构.该酶全酶晶体为八聚体,只有二聚体形式具有生物学活性.多聚体形式增加了氢键和盐桥数量,加上蛋白中含有较多的脯氨酸残基,使得该蛋白的热稳定性大大增加,CrS的晶体结构如图5所示.
5 六价铬细菌还原在污染治理中的应用
汤洁等[86]利用铁屑和Escherichia coli FR-2协同还原污染水体中的Cr(Ⅵ)取得了很好的效果; 谢翼飞等[87]将5株SRB复合菌生成的生物硫铁纳米材料用于高浓度含铬废水的处理,可在10 min内使Cr(Ⅵ)浓度0.03 mol ·L-1的废水达标排放; Kathiravan等[88]研究了Bacillus sp.在铬污染土壤生物修复中的作用,序批式和连续实验均发现土壤中Cr(Ⅵ)生物还原的效果明显; Jeyasingh等[89]采用生物屏障和反应区技术来修复铬污染地下水,已达中试规模,其主要原理是利用富集的土著微生物形成反应区域还原Cr(Ⅵ);柴立元等[90]采用从铬渣污染土壤中筛选的土著微生物Pannonibacter phragmitetus进行铬污染土壤修复,结果显示该微生物可在316 h内完全还原污染土壤中的Cr(Ⅵ)(360 mg ·kg-1); 马泽民[91]以Achromobacter sp. CH-1为基础,采用“铬渣造粒-细菌堆浸”进行了现场中试研究,运行了20 t ·批-1规模的细菌解毒铬渣处理工程,经10 d运行后,达到国家危险废物浸出毒性鉴别标准(GB 5085.3-2007); 笔者实验室采用从锰矿采冶区分离的Achromobacter sp. TY3-4对某铬渣堆场残留铬渣进行解毒,其Cr(Ⅵ)还原效果相比文献中同属菌株更好(结果待发表).从上述研究可以看到,六价铬细菌还原在污染治理领域已得到了初步的应用,显示了其巨大的潜力,但同时也存在现场还原效率较实验室低、 菌种适应能力较差的问题,因此只有对Cr(Ⅵ)细菌还原的分子机制进行深入研究才能逐步解决这些问题. 6 存在的问题及未来的研究方向
尽管已有上百种细菌被证实具有Cr(Ⅵ)还原能力,并对其特性和还原Cr(Ⅵ)的分子机制进行了许多研究,但是,还有很多科学问题有待解决:①在还原Cr(Ⅵ)为Cr(Ⅲ)的电子传递过程中,会生成中间态的Cr(Ⅴ)和Cr(Ⅳ),这种中间态的Cr是如何进行电子传递的机制尚未知晓; ②研究发现许多细菌是在体内进行Cr(Ⅵ)还原,并在体内形成Cr(Ⅲ)沉淀,这说明细胞能够将Cr(Ⅵ)大量地“吸入”体内,这使得很多研究只根据反应体系中水溶液的Cr(Ⅵ)浓度的降低来衡量其还原能力的方法有待商榷,因此需要一个严谨的检测方式来准确评估其还原能力; ③许多文献仅仅是对细胞的粗提蛋白进行了Cr(Ⅵ)还原能力的评估,纯化蛋白的机制研究极为缺乏,目前仅有少数几种蛋白进行了结构层面的解析; ④Cr(Ⅵ)直接还原膜相关的蛋白,目前仅仅是一些定性的研究,缺乏深入研究; ⑤实际环境中细菌还原Cr(Ⅵ)是一个群体行为,而不是单独一个细胞在进行,该过程细胞间的相互作用及整个细菌群体的反应鲜有文献进行深入报道; ⑥目前的研究对胞外多聚物(EPS)还原Cr(Ⅵ)关注极少,但是已有文献证明EPS确实具有该种功能[92],继续研究EPS中关键产物的基因调控和表达可以全面的解释细菌Cr(Ⅵ)的还原机制.
根据Cr(Ⅵ)细菌还原分子机制的研究现状及实验技术的发展水平,笔者认为未来该领域的发展方向主要集中在:①Cr(Ⅵ) Cr(Ⅲ)还原过程中间态Cr与相关蛋白之间结合反应的分子机制; ②Cr(Ⅵ)还原膜相关蛋白的结构与功能研究; ③Cr(Ⅵ)还原相关基因簇及其调控表达机制的研究; ④EPS中Cr(Ⅵ)还原相关分泌产物的基因表达和调控; ⑤Cr(Ⅵ)还原群体感应信号分子的基因表达和调控.虽然目前的研究对于完全解释Cr(Ⅵ)细菌还原的分子机制还有很远的距离,但通过日益发展的实验手段和技术,以及对Cr(Ⅵ)还原理论体系的逐步构建,对于该种机制的研究将会不断深入,最终揭示这种奇妙的微生物行为.
| [1] | Subhajit M, Smita R, Suresh G. Biodegradation kinetics of Cr(Ⅵ) by acclimated mixed culture[J]. Research Journal of Chemistry and Environment, 2011, 15 (2): 181-184. |
| [2] | Narayani M, Shetty K V. Chromium-resistant bacteria and their environmental condition for hexavalent chromium removal: a review[J]. Critical Reviews in Environmental Science and Technology, 2013, 43 (9): 955-1009. |
| [3] | Cheung K H, Gu J D. Mechanism of hexavalent chromium detoxification by microorganisms and bioremediation application potential: A review[J]. International Biodeterioration & Biodegradation, 2007, 59 (1): 8-15. |
| [4] | Nickens K P, Patierno S R, Ceryak S. Chromium genotoxicity: a double-edged sword[J]. Chemico-Biological Interactions, 2010, 188 (2): 276-288. |
| [5] | Romanenko V I, Korenkov V N. Pure culture of bacteria using chromates and bichromates as hydrogen acceptors during development under anaerobic conditions[J]. Microbiology, 1977, 46 (3): 414-417. |
| [6] | Barrera-Diaz C E, Lugo-Lugo V, Bilyeu B. A review of chemical, electrochemical and biological methods for aqueous Cr(Ⅵ) reduction[J]. Journal of Hazardous Materials, 2012, 223-224 : 1-12. |
| [7] | 高小朋, 张欠欠, 许平, 等. 微生物还原Cr(Ⅵ)的研究进展[J]. 微生物学通报, 2008, 35 (5): 820-824. |
| [8] | Cummings D E, Fendorf S, Singh N, et al. Reduction of Cr(Ⅵ) under acidic conditions by the facultative Fe(Ⅲ)-reducing bacterium Acidiphilium cryptum[J]. Environmental Science & Technology, 2007, 41 (1): 146-152. |
| [9] | Llovera S, Bonet R, Simonpujol M D, et al. Chromate reduction by resting cells of agrobacterium radiobacter Eps-916[J]. Applied and Environmental Microbiology, 1993, 59 (10): 3516-3518. |
| [10] | Jin H J, Zhang Y F, Buchko G W, et al. Structure determination and functional analysis of a chromate reductase from Gluconacetobacter hansenii[J]. Plos One, 2012, 7 (8): e42432. |
| [11] | Sultan S, Hasnain S. Biological chromium (Ⅵ) reduction by chromium-resistant Ochrobactrum anthropi isolated from tannery effluent[J]. New Biotechnology, 2009, 25 : S370-S370. |
| [12] | Morais P V, Branco R, Francisco R. Chromium resistance strategies and toxicity: what makes Ochrobactrum tritici 5bvl1 a strain highly resistant[J]. Biometals, 2011, 24 (3): 401-410. |
| [13] | Shi Y, Chai L Y, Yang Z H, et al. Identification and hexavalent chromium reduction characteristics of Pannonibacter phragmitetus[J]. Bioprocess and Biosystems Engineering, 2012, 35 (5): 843-850. |
| [14] | Xu L, Luo M F, Jiang C Y, et al. In vitro reduction of hexavalent chromium by cytoplasmic fractions of pannonibacter phragmitetus LSSE-09 under aerobic and anaerobic conditions[J]. Applied Biochemistry and Biotechnology, 2012, 166 (4): 933-941. |
| [15] | Sedlá Dček V, Ku Dčera I. Chromate reductase activity of the Paracoccus denitrificans ferric reductase B (FerB) protein and its physiological relevance[J]. Archives of Microbiology, 2010, 192 (11): 919-926. |
| [16] | Nepple B B, Kessi J, Bachofen R. Chromate reduction by Rhodobacter sphaeroides[J]. Journal of Industrial Microbiology & Biotechnology, 2000, 25 (4): 198-203. |
| [17] | 龙腾发. 还原Cr(Ⅵ)的特异功能菌的选育及其解毒铬的初步研究[D]. 长沙: 中南大学, 2005. 21-40. |
| [18] | Wani R, Kodam K M, Gawai K R, et al. Chromate reduction by Burkholderia cepacia MCMB-821, isolated from the pristine habitat of alkaline crater lake[J]. Applied Microbiology and Biotechnology, 2007, 75 (3): 627-632. |
| [19] | Caravelli A H, Giannuzzi L, Zaritzky N E. Reduction of hexavalent chromium by Sphaerotilus natans a filamentous micro-organism present in activated sludges[J]. Journal of Hazardous Materials, 2008, 156 (1-3): 214-222. |
| [20] | Cabrera G, Viera M, Gómez J M, et al. Bacterial removal of chromium (Ⅵ) and (Ⅲ) in a continuous system[J]. Biodegradation, 2007, 18 (4): 505-513. |
| [21] | Narayani M, Shetty V K. Characteristics of a novel Acinetobacter sp. and its kinetics in hexavalent chromium bioreduction[J]. Journal of Microbiology and Biotechnology, 2012, 22 (5): 690-698. |
| [22] | Kasatkina T, Podgorsky V, Mikhalovsky S, et al. Biotransformation of oxidised anions by selected bacteria[J]. Process Metallurgy, 1999, 9 : 673-680. |
| [23] | Komori K, Wang P C, Toda K, et al. Factors affecting chromate reduction in enterobacter-cloacae strain Ho1[J]. Applied Microbiology and Biotechnology, 1989, 31 (5): 567-570. |
| [24] | Liu G F, Yang H, Wang J, et al. Enhanced chromate reduction by resting Escherichia coli cells in the presence of quinone redox mediators[J]. Bioresource Technology, 2010, 101 (21): 8127-8131. |
| [25] | Francis C A, Obraztsova A Y, Tebo B M. Dissimilatory metal reduction by the facultative anaerobe Pantoea agglomerans SP1[J]. Applied and Environmental Microbiology, 2000, 66 (2): 543-548. |
| [26] | Suzuki T, Miyata N, Horitsu H, et al. Nad(P)H-dependent chromium(Ⅵ) reductase of Pseudomonas ambigua G-1-a Cr(Ⅴ) intermediate is formed during the reduction of Cr(Ⅵ) to Cr(Ⅲ)[J]. Journal of Bacteriology, 1992, 174 (16): 5340-5345. |
| [27] | Garg S K, Tripathi M, Singh S K, et al. Pentachlorophenol dechlorination and simultaneous Cr6+ reduction by Pseudomonas putida SKG-1 MTCC (10510): characterization of PCP dechlorination products, bacterial structure, and functional groups[J]. Environmental Science and Pollution Research, 2013, 20 (4): 2288-2304. |
| [28] | McLean J S, Beveridge T J, Phipps D. Isolation and characterization of a chromium-reducing bacterium from a chromated copper arsenate-contaminated site[J]. Environmental Microbiology, 2000, 2 (6): 611-619. |
| [29] | Myers C R, Carstens B P, Antholine W E, et al. Chromium(Ⅵ) reductase activity is associated with the cytoplasmic membrane of anaerobically grown Shewanella putrefaciens MR-1[J]. Journal of Applied Microbiology, 2000, 88 (1): 98-106. |
| [30] | Middleton S S, Bencheikh-Latmani R, Mackey M R, et al. Cometabolism of Cr(Ⅵ) by Shewanella oneidensis MR-1 produces cell-associated reduced chromium and inhibits growth[J]. Biotechnology and Bioengineering, 2003, 83 (6): 627-637. |
| [31] | Alam M Z, Ahmad S. Toxic chromate reduction by resistant and sensitive bacteria isolated from tannery effluent contaminated soil[J]. Annals of Microbiology, 2012, 62 (1): 113-121. |
| [32] | Kwak Y H, Lee D S, Kim H B. Vibrio harveyi nitroreductase is also a chromate reductase[J]. Applied and Environmental Microbiology, 2003, 69 (8): 4390-4395. |
| [33] | Chardin B, Dolla A, Chaspoul F, et al. Bioremediation of chromate: thermodynamic analysis of the effects of Cr(Ⅵ) on sulfate-reducing bacteria[J]. Applied Microbiology and Biotechnology, 2002, 60 (3): 352-360. |
| [34] | Tucker M D, Barton L L, Thomson B M. Reduction of Cr, Mo, Se and U by Desulfovibrio desulfuricans immobilized in polyacrylamide gels[J]. Journal of Industrial Microbiology & Biotechnology, 1998, 20 (1): 13-19. |
| [35] | Goulhen F, Gloter A, Guyot F, et al. Cr(Ⅵ) detoxification by Desulfovibrio vulgaris strain Hildenborough: microbe-metal interactions studies[J]. Applied Microbiology and Biotechnology, 2006, 71 (6): 892-897. |
| [36] | Lovley D R, Giovannoni S J, White D C, et al. Geobacter metallireducens gen-nov sp-nov, a microorganism capable of coupling the complete oxidation of organic-compounds to the reduction of iron and other metals[J]. Archives of Microbiology, 1993, 159 (4): 336-344. |
| [37] | Tuo Y, Liu G F, Zhou J T, et al. Microbial formation of palladium nanoparticles by Geobacter sulfurreducens for chromate reduction[J]. Bioresource Technology, 2013, 133 : 606-611. |
| [38] | Ibrahim A S S, El-Tayeb M A, Elbadawi Y B, et al. Hexavalent chromate reduction by alkaliphilic Amphibacillus sp. KSUCr3 is mediated by copper-dependent membrane-associated Cr(Ⅵ) reductase[J]. Extremophiles, 2012, 16 (4): 659-668. |
| [39] | Paul S C, Jain P, Mitra J, et al. Induction of Cr(Ⅵ) reduction activity in an Anoxybacillus strain under heat stress: a biochemical and proteomic study[J]. FEMS Microbiology Letters, 2012, 331 (1): 70-80. |
| [40] | Cheung K H, Lai H Y, Gu J D. Membrane-associated hexavalent chromium reductase of Bacillus megatelium TKW3 with induced expression[J]. Journal of Microbiology and Biotechnology, 2006, 16 (6): 855-862. |
| [41] | Cheung K H, Gu J D. Chromate reduction by Bacillus megaterium TKW3 isolated from marine sediments[J]. World Journal of Microbiology & Biotechnology, 2005, 21 (3): 213-219. |
| [42] | Zhao C Q, Yang Q H, Chen W Y, et al. Removal of hexavalent chromium in tannery wastewater by Bacillus cereus[J]. Canadian Journal of Microbiology, 2012, 58 (1): 23-28. |
| [43] | 肖伟, 王磊, 李倬锴, 等. 六价铬还原细菌Bacillus cereus S5.4还原机理及酶学性质研究[J]. 环境科学, 2008, 29 (3): 751-755. |
| [44] | Morokutti A, Lyskowski A, Sollner S, et al. Structure and function of YcnD from Bacillus subtilis, a flavin-containing oxidoreductase[J]. Biochemistry, 2005, 44 (42): 13724-13733. |
| [45] | Pal A, Dutta S, Paul A K. Reduction of hexavalent chromium by cell-free extract of Bacillus sphaericus AND 303 isolated from serpentine soil[J]. Current Microbiology, 2005, 51 (5): 327-330. |
| [46] | Sayel H, Bahafid W, Joutey N T, et al. Cr(Ⅵ) reduction by Enterococcus gallinarum isolated from tannery waste-contaminated soil[J]. Annals of Microbiology, 2012, 62 (3): 1269-1277. |
| [47] | Alam M Z, Malik A. Chromate resistance, transport and bioreduction by Exiguobacterium sp. ZM-2 isolated from agricultural soil irrigated with tannery effluent[J]. Journal of Basic Microbiology, 2008, 48 (5): 416-420. |
| [48] | Sagar S, Dwivedi A, Yadav S, et al. Hexavalent chromium reduction and plant growth promotion by Staphylococcus arlettae Strain Cr11[J]. Chemosphere, 2012, 86 (8): 847-852. |
| [49] | Nguema P F, Luo Z J. Aerobic chromium(Ⅵ) reduction by chromium-resistant bacteria isolated from activated sludge[J]. Annals of Microbiology, 2012, 62 (1): 41-47. |
| [50] | Tebo B M, Obraztsova A Y. Sulfate-reducing bacterium grows with Cr(Ⅵ), U(Ⅵ), Mn(Ⅳ), and Fe(Ⅲ) as electron acceptors[J]. Fems Microbiology Letters, 1998, 162 (1): 193-198. |
| [51] | Roh Y, Liu S V, Li G, et al. Isolation and characterization of metal-reducing thermoanaerobacter strains from deep subsurface environments of the Piceance Basin, Colorado[J]. Applied and Environmental Microbiology, 2002, 68 (12): 6013-6020. |
| [52] | Camargo F A O, Bento F M, Okeke B C, et al. Hexavalent chromium reduction by an actinomycete, Arthrobacter crystallopoietes ES 32[J]. Biological Trace Element Research, 2004, 97 (2): 183-194. |
| [53] | Asatiani N V, Abuladze M K, Kartvelishvili T M, et al. Effect of Chromium(Ⅵ) action on Arthrobacter oxydans[J]. Current Microbiology, 2004, 49 (5): 321-326. |
| [54] | Elangovan R, Philip L, Chandraraj K. Hexavalent chromium reduction by free and immobilized cell-free extract of Arthrobacter rhombi-RE[J]. Applied Biochemistry and Biotechnology, 2010, 160 (1): 81-97. |
| [55] | Ng T W, Cai Q H, Wong C K, et al. Simultaneous chromate reduction and azo dye decolourization by Brevibacterium casei: azo dye as electron donor for chromate reduction[J]. Journal of Hazardous Materials, 2010, 182 (1-3): 792-800. |
| [56] | Sani R K, Peyton B M, Smith W A, et al. Dissimilatory reduction of Cr(Ⅵ), Fe(Ⅲ), and U(Ⅵ) by Cellulomonas isolates[J]. Applied Microbiology and Biotechnology, 2002, 60 (1-2): 192-199. |
| [57] | Chatterjee S, Sau G B, Mukherjee S K. Bioremediation of Cr(Ⅵ) from chromium-contaminated wastewater by free and immobilized cells of Cellulosimicrobium cellulans KUCr3[J]. Bioremediation Journal, 2011, 15 (3): 173-180. |
| [58] | Chatterjee S, Sau G B, Mukherjee S K. Plant growth promotion by a hexavalent chromium reducing bacterial strain, Cellulosimicrobium cellulans KUCr3[J]. World Journal of Microbiology & Biotechnology, 2009, 25 (10): 1829-1836. |
| [59] | Ge S M, Zhou M H, Dong X J, et al. Distinct and effective biotransformation of hexavalent chromium by a novel isolate under aerobic growth followed by facultative anaerobic incubation[J]. Applied Microbiology and Biotechnology, 2013, 97 (5): 2131-2137. |
| [60] | Sarangi A, Krishnan C. Comparison of in vitro Cr(Ⅵ) reduction by CFEs of chromate resistant bacteria isolated from chromate contaminated soil[J]. Bioresource Technology, 2008, 99 (10): 4130-4137. |
| [61] | Amoozegar M A, Ghasemi A, Razavi M R, et al. Evaluation of hexavalent chromium reduction by chromate-resistant moderately halophile, Nesterenkonia sp. strain MF2[J]. Process Biochemistry, 2007, 42 (10): 1475-1479. |
| [62] | Poopal A C, Laxman R S. Studies on biological reduction of chromate by Streptomyces griseus[J]. Journal of Hazardous Materials, 2009, 169 (1-3): 539-545. |
| [63] | Fredrickson J K, Kostandarithes H M, Li S W, et al. Reduction of Fe(Ⅲ), Cr(Ⅵ), U(Ⅵ), and Tc(Ⅶ) by Deinococcus radiodurans R1[J]. Applied and Environmental Microbiology, 2000, 66 (5): 2006-2011. |
| [64] | Opperman D J, Van Heerden E. A membrane-associated protein with Cr(Ⅵ)-reducing activity from Thermus scotoductus SA-01[J]. Fems Microbiology Letters, 2008, 280 (2): 210-218. |
| [65] | Opperman D J, Van Heerden E. Aerobic Cr(Ⅵ) reduction by Thermus scotoductus strain SA-01[J]. Journal of Applied Microbiology, 2007, 103 (5): 1907-1913. |
| [66] | Appenroth K J, Bischoff M, Gabrys H, et al. Kinetics of chromium(V) formation and reduction in fronds of the duckweed Spirodela polyrhiza-a low frequency EPR study[J]. Journal of Inorganic Biochemistry, 2000, 78 (3): 235-242. |
| [67] | Ackerley D F, Gonzalez C F, Keyhan M, et al. Mechanism of chromate reduction by the Escherichia coli protein, NfsA, and the role of different chromate reductases in minimizing oxidative stress during chromate reduction[J]. Environmental Microbiology, 2004, 6 (8): 851-860. |
| [68] | Park C H, Gonzalez C, Ackerley D, et al. Molecular engineering of soluble bacterial proteins with chromate reductase activity[A]. In: Battelle Geneva Research Center in collaboration with AMAV, the Venetian Environmental Services Agency. Remediation and Beneficial Reuse of Contaminated Sediments[C]. Columbus: Battelle Press, 2002. 103-111. |
| [69] | Apte A D, Tare V, Bose P. Extent of oxidation of Cr(Ⅲ) to Cr(Ⅵ) under various conditions pertaining to natural environment[J]. Journal of Hazardous Materials, 2006, 128 (2-3): 164-174. |
| [70] | Richardson D J. Bacterial respiration: a flexible process for a changing environment[J]. Microbiology, 2000, 146 (Pt3): 551-571. |
| [71] | Heidelberg J F, Seshadri R, Haveman S A, et al. The genome sequence of the anaerobic, sulfate-reducing bacterium Desulfovibrio vulgaris Hildenborough[J]. Nature Biotechnology, 2004, 22 (5): 554-559. |
| [72] | 李德福, 李昕, 谢翼飞, 等. 微生物去除重金属和砷-复合硫酸盐还原菌法的机理与技术[M]. 北京: 化学工业出版社, 2011. 1-12. |
| [73] | Schicklberger M, Sturm G, Gescher J. Genomic plasticity enables a secondary electron transport pathway in Shewanella oneidensis[J]. Applied and Environmental Microbiology, 2013, 79 (4): 1150-1159. |
| [74] | Magnuson T S, Swenson M W, Paszczynski A J, et al. Proteogenomic and functional analysis of chromate reduction in Acidiphilium cryptum JF-5, an Fe(Ⅲ)-respiring acidophile[J]. Biometals, 2010, 23 (6): 1129-1138. |
| [75] | Aubert C, Lojou E, Bianco P, et al. The Desulfuromonas acetoxidans triheme cytochrome c(7) produced in Desulfovibrio desulfuricans retains its metal reductase activity[J]. Applied and Environmental Microbiology, 1998, 64 (4): 1308-1312. |
| [76] | Assfalg M, Bertini I, Bruschi M, et al. The metal reductase activity of some multiheme cytochromes c: NMR structural characterization of the reduction of chromium(Ⅵ) to chromium(Ⅲ) by cytochrome c(7)[J]. Proceedings of the National Academy of Sciences of the United States of America, 2002, 99 (15): 9750-9754. |
| [77] | Michel C, Brugna M, Aubert C, et al. Enzymatic reduction of chromate: comparative studies using sulfate reducing bacteria[J]. Applied Microbiology and Biotechnology, 2001, 55 (1): 95-100. |
| [78] | Belchik S M, Kennedy D W, Dohnalkova A C, et al. Extracellular reduction of hexavalent chromium by cytochromes MtrC and OmcA of Shewanella oneidensis MR-1[J]. Applied and Environmental Microbiology, 2011, 77 (12): 4035-4041. |
| [79] | Eswaramoorthy S, Poulain S, Hienerwadel R, et al. Crystal structure of ChrR-A quinone reductase with the capacity to reduce chromate[J]. PloS One, 2012, 7 (4): e36017. |
| [80] | Mugerfeld I, Law B A, Wickham G S, et al. A putative azoreductase gene is involved in the Shewanella oneidensis response to heavy metal stress[J]. Applied Microbiology and Biotechnology, 2009, 82 (6): 1131-1141. |
| [81] | Opperman D J, Sewell B T, Litthauer D, et al. Crystal structure of a thermostable Old Yellow Enzyme from Thermus scotoductus SA-01[J]. Biochemical and Biophysical Research Communications, 2010, 393 (3): 426-431. |
| [82] | Li X, Krumholz L R. Thioredoxin is involved in U(Ⅵ) and Cr(Ⅵ) reduction in Desulfovibrio desulfuricans G20[J]. Journal of Bacteriology, 2009, 191 (15): 4924-4933. |
| [83] | Barak Y, Ackerley D F, Dodge C J, et al. Analysis of novel soluble chromate and uranyl reductases and generation of an improved enzyme by directed evolution[J]. Applied and Environmental Microbiology, 2006, 72 (11): 7074-7082. |
| [84] | Ackerley D F, Gonzalez C F, Park C H, et al. Chromate-reducing properties of soluble Flavoproteins from Pseudomonas putida and Escherichia coli[J]. Applied and Environmental Microbiology, 2004, 70 (2): 873-882. |
| [85] | Opperman D J, Piater L A, van Heerden E. A novel chromate reductase from Thermus scotoductus SA-01 related to old yellow enzyme[J]. Journal of Bacteriology, 2008, 190 (8): 3076-3082. |
| [86] | 汤洁, 王卓行, 徐新华. 铁屑-微生物协同还原去除水体中Cr(Ⅵ)研究[J]. 环境科学, 2013, 34 (7): 2650-2657. |
| [87] | 谢翼飞, 李旭东, 李福德. 生物硫铁纳米材料特性分析及其处理高浓度含铬废水研究[J]. 环境科学, 2009, 30 (4): 1060-1065. |
| [88] | Kathiravan M N, Karthick R, Muthukumar K. Ex situ bioremediation of Cr(Ⅵ) contaminated soil by Bacillus sp.: Batch and continuous studies[J]. Chemical Engineering Journal, 2011, 169 (1-3): 107-115. |
| [89] | Jeyasingh J, Somasundaram V, Philip L, et al. Pilot scale studies on the remediation of chromium contaminated aquifer using bio-barrier and reactive zone technologies[J]. Chemical Engineering Journal, 2011, 167 (1): 206-214. |
| [90] | 柴立元, 许友泽, 王海鹰, 等. Pannonibacter phragmitetus对Cr(Ⅵ)污染土壤的修复效应[J]. 中国有色金属学报, 2009, 19 (12): 2230-2236. |
| [91] | 马泽民. Achromobacter sp. CH-1菌解毒铬渣的研究[D]. 长沙: 中南大学, 2009. 20-114. |
| [92] | Ozturk S, Aslim B, Ugur A. Chromium(Ⅵ) resistance and extracellular polysaccharide (EPS) synthesis by Pseudomonas, Stenotrophomonas and Methylobacteriumstrains[J]. Isij International, 2008, 48 (11): 1654-1658. |