 2014, Vol. 35
2014, Vol. 35


2. 武汉理工大学土木工程与建筑学院,武汉 430070
2. School of Civil Engineering and Architecture, Wuhan University of Technology, Wuhan 430070, China
硝酸盐污染不仅是造成水体富营养化的主要原因之一,而且会使人类健康受到严重威胁.若饮用水中硝酸盐浓度过高会引发高铁血红蛋白血症、 “蓝色婴儿”症状以及导致癌症.因此去除水中硝酸盐污染是目前国内外广泛关注的课题[1,2,3].一般认为生物去除硝酸氮的过程是异养反硝化细菌的代谢作用,这一代谢活动利用水中的有机物作电子供体,将硝酸盐还原成N2[4,5,6],故而需要待处理污水具有较高的COD浓度[7].但近来的一些研究发现,部分反硝化细菌可以利用铁或亚铁离子等还原态的铁基质,代替有机物还原硝酸氮.例如,Benz等[8]从含盐水沉淀中分离出1株命名为HidR2的革兰氏阴性菌种,能在pH=7.2,温度为30℃,厌氧条件下,以NO-3为电子受体把Fe2+氧化为Fe3+,并从此过程中获取能量. Weber等[9]在淡水湖泊的沉积物中筛选出1株与Pseudogulbenkiania subflava相似性达99.3%的菌种,能将硝酸盐完全地还原成气态氮产物.Shelobolina等[10]从地下蒙脱石层筛选出1株能参与铁的氧化还原反应的细菌Desulfitobacterium frappieri,它能在氧化Fe2+的同时还原硝酸盐,其最终还原产物主要为N2O.Hafenbradl等[11]从浅海海底的热水中分离出1株新的厌氧球菌Ferroglobus placidu,能以H2、 Fe2+等多种还原性物质作为电子供体进行反硝化.诸如此类微生物还包括,Chakraborty等[12]从小溪的沉积物中筛选出的Acidovorax sp.,Sorokina等[13]发现的Hoeflea siderophila,国内薛蕾等[14]报道的从厌氧污泥中分离得到1株铁自养厌氧反硝化细菌Pseudomonas stutzeri等.与传统的异养反硝化细菌相比,这类细菌可以利用无机铁基质取代有机物作为电子供体,因此在工艺上有降低运行成本的可能[15, 16].
笔者从武汉市东湖深层底泥中筛选出1株铁基质自养反硝化细菌,命名为W5.对其最佳脱氮生长条件和反硝化特性进行了研究,以期为铁自养反硝化细菌应用于工程实践提供理论依据,为开发新型生物脱氮工艺提供一定的技术支持.
菌种取自武汉市东湖深层底泥.
铁基质自养反硝化培养基(g ·L-1):FeSO4 ·7H2 O 1.8; KNO3 0.25; MgSO4 0.2; CaCl2 0.2; K2HPO4 0.3; KH2PO4 0.5; NaCl 1; NaHCO3 2; pH 6.8~7.5[16].
1.2 细菌的分离纯化从东湖深层底泥悬浊液中取30 mL样品加入100 mL灭菌后的铁基质自养反硝化密闭培养基中,于30℃下在恒温振荡器上振荡48 h后,取30 mL菌悬液接种于新的100 mL铁基质自养反硝化培养基中,30℃摇床富集培养多次,待菌悬液中的硝氮有一定的去除率后,将富集培养液接种于培养基的固体平板上,于生化培养箱30℃下培养1周,待形成菌落后挑取单菌落,在平板上划线分离.为保证所筛菌株的纯度,可重复平板划线分离2~3次.
1.3 菌株的生理生化鉴定菌株生理生化特性的鉴定参照文献[17]进行. 1.4 16S rRNA基因的PCR扩增、 序列测定
以基因组DNA为模板扩增16S rRNA基因,扩增采用的引物是一对通用引物[18].引物由上海生工生物工程有限公司合成.PCR反应体系(50 μL):10×PCR缓冲液5 μL,MgCl2(25mmol ·L-1)4 μL,dNTP 2 μL,引物BSF8/20和引物BSR1541/20各1 μL,模板DNA 1 μL, Taq酶(10000 U ·mL-1)0.5 μL,重蒸水35.5 μL. PCR程序如下; 94℃ 2 min; 94℃ 1 min,56℃ 1 min,72℃ 2 min; 第2步循环29次; 72℃ 10 min; 60℃ 10 min.PCR产物的纯化和测序由上海生工生物工程有限公司完成.
1.5 16S rRNA的系统发育分析将测序结果在GenBank数据库中进行BLAST(http://blast.ncbi.nlm.nih.gov/)序列的相关性搜索,同时利用相关种属的16S rRNA基因序列,构建系统发育树.序列对排用CLUSTAL X1.8[19]进行多序列匹配排列,进化树的构建用Neighbour-joining方法[20].进化树分枝模式的稳定性用MEGA4软件进行Bootstrap分析,重复500次,计算各分支的支持度,最后采用NJ程序[21]构建基于16S rRNA基因序列的系统发育树.
1.6 菌株最适脱氮条件及脱氮特性通过间歇试验对菌株W5的最适脱氮条件加以确定,主要考察不同进水硝酸氮浓度、 Fe2+浓度、 pH值对菌株自养反硝化效果的影响,并对其最佳培养条件下的脱氮特性进行研究.以5%的接菌量接种于密闭的摇瓶中,30℃下培养7 d后测定培养液中氨氮、 亚硝态氮、 硝态氮及Fe2+含量.
1.7 分析方法NO-2-N:N-(1-萘基)-乙二胺光度法; NO-3-N:麝香草酚分光光度法; NH+4-N:采用纳氏试剂分光光度法测定; Fe2+:邻菲罗啉分光光度法; pH 值:由320型pH 计测定; 氮气(N2):Agilent HP4890D型气相色谱仪,气相色谱的色谱柱编号GS-03-106型,条件为:载气流量He 20 mL ·min-1,柱前压0.08 MPa,柱温60℃,检测器温度120℃,气化室温度120℃; 细菌计数采用吖啶橙染色计数法.
通过富集培养,分离筛选得到1株铁基质自养反硝化菌,命名为W5,其原子力显微镜照片如图 1所示.可以看出,菌株W5呈杆状.其菌株形成的菌落呈现白色,圆形,表面和边缘光滑,菌落较分散.W5菌株的生理生化试验结果见表1.对其进行纯培养可得其生长曲线如图 2.
 | 图 1 菌株W5原子力照片Fig. 1 AFM micrograph of cells of strain W5 |
 | 图 2 菌株W5生长曲线Fig. 2 Growth curve of strain W5 |
经PCR扩增与测序,获得该菌株的部分16S rRNA基因序列,将所测菌株的序列通过BLAST检索与GenBank中的核酸序列进行比对,结果显示菌株W5的16S rRNA序列与微杆菌属(Microbacterium)具有高度同源性.利用CLUSTALX和MEGA软件,以Neighbor-Joining法绘制16S rRNA系统发育树,如图 3所示. 从中可见,菌株W5与Microbacterium oxydans strain JN270 (KF150504.1)的亲缘关系最接近,具有99%的同源性.结合菌株W5的生理生化鉴定结果,可确定菌株W5属于微杆菌属(Microbacterium sp.).
 | 图 3 菌株W5基于16S rRNA基因序列NJ法系统发育树分析 Fig. 3 Neighbor-joining phylogenesis tree based on 16S rRNA sequence of W5 and close strain |
在初始pH=7.0,温度30℃,Fe2+浓度500 mg ·L-1条件下,调整初始NO-3-N底物浓度分别为20、 30、 40、 50、 60 mg ·L-1,考察了菌株W5在不同初始硝酸氮浓度下的自养反硝化效果.经过7 d培养后,测定培养液中NO-3-N和NO-2-N的含量.
| 表1 菌株的生理生化特性1) Table 1 Physiological and biochemical characteristics of strain W5 |
从图 4可以看出,菌株W5在底物NO-3-N浓度小于40 mg ·L-1时,硝酸氮去除率随着底物NO-3-N浓度的增加而缓慢上升,当底物浓度为40 mg ·L-1时,菌株W5对硝酸氮去除率达到最高值为86.03%.进一步增加NO-3-N浓度,反硝化效果出现降低趋势.这是因为在底物NO-3-N浓度较低时,W5菌株可利用的NO-3-N浓度不足,底物NO-3-N是W5菌株代谢活动的限制因素,W5菌株的活性随着底物NO-3-N浓度的增加而增强.当NO-3-N达到40 mg ·L-1,底物NO-3-N充足,大于W5代谢活动所需的最高含量,此时NO-3-N已不再是反硝化代谢的限制因素,继续提高其含量并不能使菌株W5对NO-3-N的利用率增加.此时所去除的NO-3-N含量基本不变,而底物NO-3-N浓度增加,故而呈现出硝酸氮去除率降低的趋势.可见,菌株W5的初始最适NO-3-N浓度为40 mg ·L-1.在整个培养过程中,W5菌株的培养液中NO-2-N含量保持在极低的水平,其值为0.15~0.23 mg ·L-1.
 | 图 4 不同NO-3-N底物浓度对硝酸氮去除率的影响 Fig. 4 Effect of NO-3-N on nitrate nitrogen removal rate |
控制初始pH=7.0,温度为30℃,菌株W5的初始NO-3-N浓度为40 mg ·L-1,调整初始Fe2+浓度分别为300、 400、 500、 600、 700 mg ·L-1,经过7 d培养后测定培养液中NO-3-N和NO-2-N的含量,考察不同Fe2+浓度对菌株W5自养反硝化效果的影响.结果如图 5所示.
 | 图 5 不同Fe2+底物浓度对硝酸氮去除率的影响 Fig. 5 Effect of Fe2+ on nitrate nitrogen removal rate |
由图 5可知,菌株W5在底物Fe2+浓度小于500 mg ·L-1时,其对硝氮的去除率随着Fe2+浓度的增加而上升,当Fe2+浓度达到500 mg ·L-1时,硝氮去除率达到85.72%.继续增加Fe2+浓度,硝氮去除率随之降低.这是因为在底物Fe2+浓度较低时,底物Fe2+也是菌株W5代谢活动的限制因素,菌株W5的活性随着Fe2+浓度的增加而增强.当Fe2+到达500 mg ·L-1,铁基质形成的水合物开始大量附着在微生物细胞壁上,若继续增加Fe2+浓度会阻碍微生物与外界环境的物质交换,使得硝酸氮去除率降低.可见,菌株W5在的Fe2+浓度为500 mg ·L-1时可获得最佳的自养反硝化脱氮效果.在整个培养过程中,W5菌株的培养液中NO-2-N含量保持在极低的水平,其值为0.17~0.36 mg ·L-1.
2.5 pH值对铁自养反硝化的影响在初始温度为30℃,菌株W5的初始NO-3-N含量分别为40 mg ·L-1,初始Fe2+浓度为500 mg ·L-1的条件下,通过调节培养液初始pH值分别为5.5、 6.2、 6.8、 7.0、 8.0、 9.0,来考察初始pH值对菌株W5的铁自养反硝化效果的影响.经过7 d的培养后,测定培养液中NO-3-N、 NO-2-N、 NH+4-N和Fe2+的含量.结果如图 6所示.
 | 图 6 不同pH对硝酸氮去除率的影响Fig. 6 Effect of pH on nitrate nitrogen removal rate |
从图 6中可以看出,菌株W5在中性及偏弱酸性环境时,表现出较好的自养反硝化效果.当pH值为7.0时,菌株W5对硝酸氮的去除率为83.80%.当pH值为6.8时,菌株W5对硝酸氮的去除率达到86.35%.当pH值为9或5.5时,反硝化效果较中性偏弱酸性条件明显下降,W5菌株对硝酸氮的去除率较pH值为6.8时分别下降了13.35%和16.35%.在整个培养过程中,未出现NO-2-N的累积,培养液中NO-2-N的含量为0.08~0.51 mg ·L-1.
菌株W5在进行自养反硝化脱氮时,在将Fe2+氧化为Fe3+的同时,可将NO-3-N还原为N2,反应方程式(1)如下:
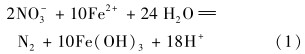
可见,铁自养反硝化过程所产生的H+会降低环境的pH值.pH值对细菌的影响是改变底物和菌体酶蛋白的带电状态[22],从而影响其对营养物质的吸收以及代谢过程中酶的活性.一些研究者认为铁自养反硝化的最适pH为中性左右[23, 24].本试验中以Fe2+作为电子供体降解NO-3-N的最佳pH值与文献报道的基本一致.当初始pH值为酸性或碱性的环境时,反硝化效果下降,主要是因为菌株在酸性或碱性条件下硝酸盐还原酶的活性受到抑制,所以造成对NO-3-N降解能力的下降.
2.6 菌株在最适条件下的反硝化特性通过对菌株W5的影响因素分析,获得了菌株W5的最适脱氮条件.在此条件下,通过连续监测培养液中NO-3-N、 NO-2-N、 NH+4-N、 N2和Fe2+的含量,研究菌株W5的自养反硝化特性,结果如图 7所示.
 | 图 7 菌株W5最佳脱氮条件下的反硝化特性 Fig. 7 Denitrification performance of strain W5 in the optimized condition |
从图 7可以看出,在7 d厌氧培养条件下,NO-3-N浓度快速下降.W5菌株培养液中NO-3-N浓度从初始的40.1 mg ·L-1降到 5.2 mg ·L-1,硝酸氮去除率可达87.0%,反硝化速率最高可达20.5 mg ·(L ·d)-1.而Godini等[25]测得异养反硝化的速率(以NO-3-N计)为740 mg ·(L ·d)-1,可见铁基质自养反硝化在生物处理系统中并非主流和有效的脱氮途径.在整个培养过程中未见亚硝酸盐的积累.培养液中NO-2-N 平均含量为0.23 mg ·L-1,其最高值不超过0.31 mg ·L-1.此外,在菌株W5的反硝化过程中,没有检测出NH+4-N的生成.同时结合培养瓶内所生成氮气含量,可以确定培养液中的硝酸氮大部分被转化成N2.图 8显示了菌株W5在最适条件下氧化利用Fe2+的情况.从中可以看出,经过1周的培养,W5培养液中Fe2+的浓度从初始的507 mg ·L-1降到 24.3 mg ·L-1,最终Fe2+的氧化率为95.2%.
 | 图 8 菌株W5最佳脱氮条件下Fe2+变化曲线 Fig. 8 Fe2+ assumption of strain W5 in the optimized condition |
由方程式(1)理论分析,根据Fe2+实际消耗量481.7 mg ·L-1可知NO-3-N理论去除量为24.09 mg ·L-1.但实际NO-3-N去除量为34.90 mg ·L-1.这可能是菌体在生长过程中由于自身的生长代谢也会同化部分氮素,并将之贮存在细胞内的缘故.有研究指出,缺少NH+4-N的外界培养条件会显著促进微生物对NO-3-N的同化作用[26].因此,菌株W5在仅有NO-3-N作为氮源的培养基中,会将部分硝酸氮同化为菌体内的有机氮,因而得到了除反硝化之外的硝酸氮去除效果.
(1)从东湖深层底泥中获得1株铁自养反硝化菌株,经形态观察、 生理生化试验和16S rRNA测序,确定菌株W5为微杆菌属(Microbacterium sp.).
(2)菌株W5的最适脱氮培养条件:NO-3-N 40 mg ·L-1,Fe2+ 500 mg ·L-1,pH 6.8~7.0.在此条件下,W5菌株对硝酸氮的反硝化速率最高可达20.5 mg ·(L ·d)-1.在整个培养过程中亚硝酸氮的产生量很少,硝酸氮大部分被转化成N2.同时作为电子供体的Fe2+的氧化率达95.2%.
| [1] | Bae B U, Jung Y H, Han W W, et al. Improved brine recycling during nitrate removal using ion exchange[J]. Water Research, 2002, 36 (13): 3330-3340. |
| [2] | Murphy A P. Chemical removal of nitrate from water [J]. Nature, 1991, 350 (6315): 223-225. |
| [3] | Karanasios K A, Vasiliadou I A, Pavlou S, et al. Hydrogenotrophic denitrification of potable water: A review [J]. Journal of Hazardous Materials, 2010, 180 (1-3): 20-37. |
| [4] | Yang G C C, Lee H L. Chemical reduction of nitrate by nanosized iron: kinetics and pathways [J]. Water Research, 2005, 39 (5): 884-894. |
| [5] | 陈立强, 郑寿荣, 尹大强, 等. 载体特性对Pd-Cu/TiO2催化剂催化脱氮性能的影响[J]. 环境化学, 2005, 24 (5): 502-505. |
| [6] | Archna, Sharma S K, Sobti R C. Nitrate removal from ground water: A review [J]. E-Journal of Chemistry, 2012, 9 (4): 1667-1675. |
| [7] | 陆彩霞, 顾平. 氢自养反硝化去除饮用水中硝酸盐的试验研究[J]. 环境科学, 2008, 29 (3): 671-676. |
| [8] | Benz M, Brune A, Schink B. Anaerobic and aerobic oxidation of ferrous iron at neutral pH by chemoheterotrophic nitrate-reducing bacteria [J]. Archives of Microbiology, 1998, 169 (2): 159-165. |
| [9] | Weber K A, Hedrick D B, Peacock A D, et al. Physiological and taxonomic description of the novel autotrophic, metal oxidizing bacterium, Pseudogulbenkiania sp. strain 2002.[J]. Applied Microbiology and Biotechnology, 2009, 83 (3): 555-565. |
| [10] | Shelobolina E S, Van Praagh C G, Lovley D R. Use of ferric and ferrous Iron containing minerals for respiration by Desulfitobacterium frappieri [J]. Geomicrobiology Journal, 2003, 20 (2): 143-156. |
| [11] | Hafenbradl D, Keller M, Dirmeier R, et al. Ferroglobus placidus gen. nov., sp. nov., a novel Hyperthermophilic archaeum that oxidizes Fe2+ at neutral pH under anoxic conditions [J]. Archives of Microbiology, 1996, 166 (5): 308-314. |
| [12] | Chakraborty A, Roden E E, Schieber J, et al. Enhanced growth of Acidovorax sp. strain 2AN during nitrate-dependent Fe(Ⅱ) oxidation in batch and Continuous-Flow systems [J]. Applied and Environmental Microbiology, 2011, 77 (24): 8548-8556. |
| [13] | Sorokina A Y, Chernousova E Y, Dubinina G A. Hoeflea siderophila sp. nov., a new neutrophilic iron-oxidizing bacterium [J]. Microbiology, 2012, 81 (1): 59-66. |
| [14] | 薛蕾, 蒯琳萍. 一株新反硝化菌的鉴定及在废水处理中的应用[J]. 环境科学与技术, 2008, 31 (7): 98-100. |
| [15] | Nielsen J L, Nielsen P H. Microbial nitrate-dependent oxidation of ferrous iron in activated sludge[J]. Environmental Science & Technology, 1998, 32 (22): 3556-3561. |
| [16] | Straub K L, Buchholz-Cleven B E E. Enumeration and detection of anaerobic ferrous iron-oxidizing, nitrate-reducing bacteria from diverse European sediments [J]. Applied and Environmental Microbiology, 1998, 64 (12): 4846-4856. |
| [17] | 东秀珠, 蔡妙英. 常见细菌系统鉴定手册 [M]. 北京: 科学出版社, 2001. |
| [18] | Devereux R, Willis S G. Amplification of ribosomal RNA sequences. Molecular microbial ecology manual [M]. Netherland: Kluwer Academic Publishers, 1995. 1-11. |
| [19] | Thompson J D, Gibson T J, Plewniak F, et al. The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools [J]. Nucleic Acids Research, 1997, 25 (24): 4876-4882. |
| [20] | Saitou N, Nei M. The neighbor-joining method: a new method for reconstructing phylogenetic trees[J]. Molecular Biology and Evolution, 1987, 4 (4): 406-425. |
| [21] | Perrière G, Gouy M. WWW-query: an on-line retrieval system for biological sequence banks[J]. Biochimie, 1996, 78 (5): 364-369. |
| [22] | 许保玖, 龙腾锐. 当代给水与废水处理原理 [M]. (第二版). 北京: 高等教育出版社, 2000. 329-330. |
| [23] | Straub K L, Benz M, Schink B, et al. Anaerobic, nitrate-dependent microbial oxidation of ferrous iron[J]. Applied and Environmental Microbiology, 1996, 62 (4): 1458-1460. |
| [24] | Kumaraswamy R, Sjollema K, Kuenen G, et al. Nitrate-dependent [Fe(Ⅱ)EDTA]2- oxidation by Paracoccus ferrooxidans sp. nov., isolated from a denitrifying bioreactor[J]. Systematic and Applied Microbiology, 2006, 29 (4): 276-286. |
| [25] | Godini H, Rezaee A, Khavanin A, et al. Heterotrophic biological denitrification using microbial cellulose as carbon source [J]. Journal of Polymers and the Environment, 2011, 19 (1): 283-287. |
| [26] | Burger M, Jackson L E. Microbial immobilization of ammonium and nitrate in relation to ammonification and nitrification rates in organic and conventional cropping systems [J]. Soil Biology and Biochemistry, 2003, 35 (1): 29-36. |