 2014, Vol.
2014, Vol. 



2. 中国科学院水生生物研究所藻类生物学重点实验室, 武汉 430072;
3. 武汉理工大学理学院, 武汉 430070
2. Key Laboratory of Algal Biology, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan 430072, China;
3. School of Sciences, Wuhan University of Technology, Wuhan 430070, China
生物土壤结皮主要是由蓝藻优先在土表拓殖生长,表土稳定后其他真核藻类、 菌类、 地衣、 苔藓等逐渐也生长其上形成易剥离的生物土壤复合层[1]. 它们广泛存在于干旱、 半干旱地区,甚至在一些区域占到生物覆盖的70%以上[2,3]. 研究人员根据结皮发育演替的程度以及其中优势群落的不同,常把生物结皮分为藻结皮、 地衣结皮和藓结皮[4]. 藻结皮代表发育演替的初级阶段,但它们却能够通过改善表土微环境来增加晚期结皮物种拓殖生存的可能性,从而促进结皮的发育演替[5, 6, 7, 8, 9]. 因此,通过人工方法构建藻结皮从而达到促进生物结皮的发育以及土壤生态系统的恢复,从20世纪80年代就被研究者们提上了议事日程[3, 10, 11, 12, 13, 14, 15].
一些研究人员通过浇灌结皮泥浆、 接种揉碎的结皮碎末以恢复结皮的想法都已在方法上得到了验证[10,11],但由于接种物的限制使得这些技术没有被广泛使用. 同时也有人提出接种蓝藻以构建生物结皮的想法,并且在实验室逐渐得到验证[3,12]. 然而进程不仅仅停留在实验室内部,Hu等[13,14]在室内外的实验中研究发现,蓝藻结皮同样适合野外接种构建. 有了这样的研究前提,本研究小组从2002年起便在内蒙古达拉特旗(库布齐沙漠东缘)通过接种蓝藻在野外进行了生物结皮的大规模构建[16, 17, 18, 19, 20]. 在本研究区域,随着蓝藻的接种以及结皮的构建,该地区环境逐渐改善,结实的生物结皮大面积形成. 同时研究表明随着藻结皮的形成,表土吸收非降雨型水分的能力增强,持水量也明显增加[19,20]. 然而接种蓝藻形成藻结皮后,随着结皮向晚期的发育演替,表土持水特性又将发生怎样的变化?这种变化与结皮发育演替过程中土壤理化特性的变化又有何关系?这些都是人们目前亟需解决的问题. 因此本研究以达拉特旗实验区人工接种蓝藻形成的生物结皮为对象,进而探讨随着生物结皮的发育演替,表土持水特性的变化情况以及影响持水性能的环境因素. 通过研究发育演替对结皮水分保持能力的影响,以期为不同类型结皮自身生长发育演替特点的探讨和干旱半干旱地区退化生态系统的人工恢复和演替提供一定理论依据.
达拉特旗实验区位于库布齐沙漠东缘(40°21′N,109°51′E)的人工植被恢复区. 该地区海拔1 040 m,为高原荒漠与荒漠草原过渡地带,属典型的大陆性季风气候; 年平均气温6.1℃,最高气温40.2℃,最低气温为-34.5℃; 年平均降雨量为240~300 mm,一年内的雨量分布很不均匀,主要集中在7~9月; 每年生长季节(5~9月)的潜在蒸发量约为2 400 mm; 该地区的年平均风速为3.3 m ·s-1,主要为西北风,大于5 m ·s-1的起沙风每年超过180 d. 土壤为疏松贫瘠的流沙,沙丘平均相对高差为5 m[16, 17, 20]. 天然植被以沙米(Agriophyllum squarrosum)和虫实(Leymus chinensis)为主. 为了加速该地区生物结皮的形成以及生态环境的恢复,两种丝状蓝藻(具鞘微鞘藻Microcoleus vaginatus和爪哇伪枝藻Scytonema javanicum)从2002年开始陆续被混合接种于流动沙丘表面.
实验中所有样品在蓝藻接种固沙区和流沙区的5个样点进行采样,样点设计及各样点微地势见表1,其中包括不同发育演替阶段的2种生物结皮:藻结皮和藓结皮,并以流沙作为对照. 结皮及流沙样品用环刀和无菌铲采集后装于无菌培养皿中,Parafilm 膜封口,所有样品快速运回实验室后一个月内进行相关的实验分析.
| 表1 实验位点概述 Table 1 Introduction of experimental sites |
本实验中结皮的生物量用叶绿素a表示(确切是指结皮中的光合生物量). 将采取的结皮样品研磨于10 mL 95%热乙醇(80℃),在恒温水浴锅中80℃萃取2 min,之后置于4℃的冰箱中黑暗静置6 h,提取物在5 000 r ·min-1离心10 min,取上清于波长665 nm和750 nm处测定吸光值,然后加5 滴1 mol ·L-1 盐酸酸化,10 min 后再于波长665 nm 和750 nm处测定吸光值. 结果计算公式为:
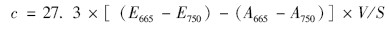
结皮含水量在样品采集运回实验室后立即开始采用烘干法(105℃,24 h)测定,结果计算为:
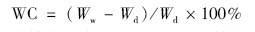
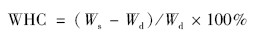
结皮相对持水量(RWC,%)计算为:
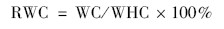
容重:环刀法[21]; 土壤质地:吸管法[21]; 多糖含量:苯酚硫酸法[22].
实验中方差分析利用One-way ANOVA进行分析,相关性分析利用Bivariate Correlations进行分析,回归分析利用Linear Regression进行分析,所有分析在SPSS 13.0软件上进行.
两种丝状蓝藻——具鞘微鞘藻(M. vaginatus)和爪哇伪枝藻(S. javanicum)通过工程化培养后按一定比例接种于流沙沙丘以加速沙表生物结皮的形成及生态环境的恢复. 在水分条件充足的情况下,接种后一个星期结皮开始形成,但刚形成的结皮很薄,颜色较浅,平坦、 易碎,易被风吹走. 随着结皮的生长发育,其厚度逐渐增加,颜色变深,在适宜的环境条件下(如水分、 表面粗糙度、 地形等),2~3 a后一些微地型开始萌发藓类,藻结皮逐渐向藓结皮演替(图 1). 与较平坦的藻结皮相比,藓结皮表面由地毯状的藓类植物所覆盖,结皮更加厚实,表面更加粗糙; 藻结皮一般为灰白色或深灰色,藓结皮为褐色或墨绿色; 而且随着结皮的演替,结皮中的优势种也由藻结皮中的藻类(如具鞘微鞘藻)演替为藓结皮中的藓类.
 | 图 1 生物结皮的发育演替及其中的优势种变化 Fig.1 Development,succession and change of dominant species in biological soil crusts (a)藻结皮; (b)藻结皮中的优势种——具鞘微鞘藻; (c)藓结皮; (d)藓结皮中的优势种 |
在研究区域,藻结皮直接演替为藓结皮,到目前为止没有地衣结皮的出现,因此,在本研究中只有藻结皮与藓结皮两种结皮类型. 从图 2可以发现结皮的出现明显增加了表土生物量,而且随着结皮的演替,藓结皮的生物量明显高于藻结皮. 同时,随着结皮的发育演替,结皮多糖含量、 厚度以及孔隙度增加,容重减少. 在土壤质地组成方面,随着结皮的形成以及发育演替,粉粒、 黏粒含量增加,沙粒逐渐减少. 与流沙相比,藻结皮中粉粒含量增加34.9~37.2倍,黏粒含量增加16.4~17.7倍; 藓结皮中粉粒含量增加65.4~67.5倍,黏粒含量增加51.2~52.9倍. 但不论结皮还是流沙,其机械组成仍是以沙粒为主,粉粒、 黏粒含量都很低,粉粒与黏粒之和一般<350 g ·kg-1(表2).
 | 图 2 不同发育演替阶段生物结皮的生物量箱形图 Fig.2 Box and whisker plots of crust biomass in the different developmental and successional biological soil crusts |
| 表2 不同实验位点生物结皮的理化特性 Table 2 Physicochemical properties of biological soil crusts in different experimental sites |
由图 3可以发现不同采样点结皮的含水量及饱和持水量都出现很大的变化,在实验中结皮及流沙持水量为0.2% ~0.5%,而饱和持水量为20% ~50%,相对持水量则比较恒定,一般在1%左右(0.87%~1.05%). 然而从图 3仍然可以发现随着结皮的演替,结皮的含水量及饱和持水量呈现规律性增加,即:藓结皮>藻结皮>流沙. 而且藻结皮中的含水量(饱和持水量)为流沙中的1.1~1.3倍,藓结皮中的含水量(饱和持水量)为流沙中的1.8~2.2倍.
 | 图 3 不同实验位点与不同发育演替阶段生物结皮的含水量与饱和持水量
Fig.3 Water holding capacity and water content of
biological soil crusts in the different experimental sites
and different developmental and successional stages
|
表3为结皮含水量及饱和持水量与结皮理化特性之间的相关分析结果,从中可以发现结皮含水量(饱和持水量)与结皮生物量、 多糖含量、 厚度、 容重、 孔隙度以及土壤质地都存在显著相关性(P<0.05),其中与结皮生物量、 多糖含量、 厚度、 容重、 粉粒和黏粒含量呈正相关,与结皮孔隙度和沙粒含量呈负相关. 以结皮含水量为因变量进行逐步回归分析时,黏粒含量进入回归模型; 而以结皮饱和持水量为因变量时,孔隙度进入回归模型,且它们的R2值均>0.98(P<0.001,表4).
| 表3 结皮含水量及饱和持水量与结皮理化特性之间的相关性分析 1)Table 3 Correlation analyses between water holding capacity,water content and crust physicochemical properties |
| 表4 结皮理化特性对其含水量及饱和持水量的逐步回归分析结果 Table 4 Stepwise regression analysis of crust water holding capacity and water content based on crust physicochemical properties |
在实验区,为了形成生物结皮,具鞘微鞘藻(M. vaginatus)和爪哇伪枝藻(S. javanicum)被混合接种于流沙. 接种后藻丝直接与沙土颗粒接触,当它们生长移动时,便会束缚沙土颗粒形成团状物[13, 14, 17, 23]; 同时,它们也向胞外分泌且积累多糖聚合物,从而更进一步胶结沙土颗粒,形成更为稳定的团聚物[8, 17, 24]. 在本研究中发现,接种一个星期之后结皮便开始形成,但此时结皮比较薄,颜色也较浅,平坦而且易碎,很容易被风吹走. 随着结皮的生长,其颜色逐渐变深,表面逐渐粗糙,厚度也越来越厚. Chen等[16]研究发现接种20 d后,结皮便可以抵御19 mm降水和4.3级风力的侵蚀. 同时,随着结皮的形成,其中的藻类不断生长繁殖,真核藻类逐渐出现. 在Wang等[22]的研究中发现,在接种3 a后结皮中出现14种藻类(包括蓝藻和真核藻类),而且藻类生物量随着结皮的生长发育增加. 之后,随着结皮的发育演替,藓类在结皮上出现,此时藻类生物量便开始下降[25]. 但当结皮发育为藓结皮后,由于大量藓类的出现,结皮的总光合生物量则进一步增加. 这与Büdel等[26]得到的实验结果一致,即随着结皮的发育演替其中叶绿素a含量呈增加的趋势.
生物结皮的发育演替与周围环境条件密切相关. 周围环境条件(如地形地势、 光照、 温度、 土壤结构、 类型以及营养条件等)在很大程度上可以影响结皮的发育演替,甚至在一些条件的限制下结皮可能完全停止发育,从而滞留于演替的某一阶段[27,28]. 在本实验区,一些微地形下的藻结皮2~3 a后逐渐向藓结皮演替,这可能表明这里的土壤微环境,如土壤质地、 水分、 营养条件被逐渐改善. 另一方面,结皮的发育演替同样可以改变周围的微环境. 在本实验中发现,随着结皮的发育演替,表土持水能力显著提高,这与表土理化特性的改变有着密切的关系. 因为随着结皮的发育演替,结皮生物量的增加以及结皮多糖的积累,结皮不断从空气中截留粉粒、 黏粒,从而使粉粒、 黏粒在结皮中不断富积,而沙粒则呈减少趋势; 同时,随着结皮厚度的增加以及粉粒、 黏粒含量的增加,结皮容重减小,孔隙度增加. 这可能是由于较小比重的粉粒、 黏粒含量的增加从而使结皮容重减小; 也可能是由于结皮生物的生长代谢(藻丝、 菌丝的运动以及代谢分泌物的积累)使得结皮层孔隙度增加.
在干旱半干旱地区由于有限的降雨以及强烈的蒸发,表土含水量一般非常有限,这就在一定程度上限制了表土微生物及其他土壤生物的生存. 生物结皮的形成发育演替,不仅改善了表土理化特性,增加了表土持水能力,而且使得表土能够更加有效地利用荒漠地区的各种水分来源,如雾、 露以及水汽的吸收[19,20],这就使得结皮中含水量明显高于流沙. 此外,本研究也发现随着结皮的发育演替,其持水能力呈增加的趋势,含水量也相应增加. 尽管相关分析得到结皮很多特性都与其含水量及饱和持水量相关,但通过逐步回归分析表明,影响结皮持水量的主要因素是结皮中黏粒的含量,而影响结皮饱和持水量的主要因素是结皮的孔隙度.
(1) 接种荒漠蓝藻有助于生物结皮的发育演替. 人工接种荒漠蓝藻之后,藻结皮能够很快形成,并在一些微环境下(一般在背风坡)藻结皮2~3 a后演替为藓结皮.
(2) 生物结皮的发育演替促进了表土理化特性的改善. 随着结皮的发育演替,表土生物量、 多糖含量、 结皮厚度以及孔隙度增加,而土壤容重减小; 同时形成结皮之后,表土质地也发生了明显的变化,其中沙粒含量逐渐减少,粉粒、 黏粒含量增加.
(3) 生物结皮的发育演替明显改善了表土持水特性,分析表明表土含水量及饱和持水量与表土生物量、 多糖含量、 结皮厚度、 容重、 粉粒和黏粒含量呈正相关,其中影响含水量的主要因素为表土黏粒的含量,而影响饱和持水量的主要因素为表土孔隙度.
| [1] | Hu C X, Zhang D L, Liu Y D. Research progress on algae of the microbial crusts in arid and semiarid regions[J]. Progress in Natural Science, 2004, 14 (4): 289-295. |
| [2] | Eldridge D J, Greene R S B. Microbiotic soil crust: a review of their roles in soil and ecological processes in the rangelands of Australia[J]. Australian Journal of Soil Research, 1994, 32 (3): 389-415. |
| [3] | Evans R D, Johansen J R. Microbiotic crusts and ecosystem processes[J]. Critical Reviews in Plant Sciences, 1999, 18 (2): 183-225. |
| [4] | Rivera-Aguilar V, Godínez-Alvarez H, Manuell-Cacheux I, et al. Physical effects of biological soil crusts on seed germination of two desert plants under laboratory conditions[J]. Journal of Arid Environments, 2005, 63 (1): 344-352. |
| [5] | Kurina L M, Vitousek P M. Controls over the accumulation and decline of a nitrogen-fixing lichen, Stereocaulon vulcani, on young Hawaiin lava flows[J]. Journal of Ecology, 1999, 87 (5): 784-799. |
| [6] | Acea M J, Prieto-Fernández A, Diz-Cid N. Cyanobacterial inoculation of heated soils: effect on microorganisms of C and N cycles and on chemical composition in soil surface[J]. Soil Biology & Biochemistry, 2003, 35 (4): 513-524. |
| [7] | 饶本强, 李华, 熊瑛, 等. 实验室条件下蓝藻结皮对低温光照胁迫的响应与微结构变化[J]. 环境科学, 2012, 33 (8): 2793-2803. |
| [8] | Hu C X, Liu Y D. Parimary succession of algal community structure in desert soil[J]. Acta Botanica Sinica, 2003, 45 (8): 917-924. |
| [9] | Langhans T M, Storm C, Schwabe A. Regeneration processes of biological soil crusts, macro-cryptogams and vascular plants pecies after fine-scale disturbance in a temperate region: Recolonization or successional replacement?[J]. Flora-Morphology, Distribution, Functional Ecology of Plants, 2010, 205 (1): 46-60. |
| [10] | St Clair L L, Johansen J R, Webb B L. Rapid stabilization of firedisturbed sites using a soil crust slurry: inoculation studies[J]. Reclamation & Revegetation Research, 1986, 4: 261-269. |
| [11] | Belnap J. Recovery rates of cryptobiotic crusts: inoculant use and assessment methods[J]. Great Basin Naturalist, 1993, 53 (1): 89-95. |
| [12] | Acea M J, Diz N, Prieto-Fernández A. Microbial populations in heated soils inoculated with cyanobacteria[J]. Biology and Fertility of Soils, 2001, 33 (2): 118-125. |
| [13] | Hu C X, Liu Y D, Zhang D L, et al. Cementing mechanism of algal crusts from desert area[J]. Chinese Science Bulletin, 2002, 47 (16): 1361-1368. |
| [14] | Hu C X, Liu Y D, Song L R, et al. Effect of desert soil algae on the stabilization of fine sands[J]. Journal of Applied Phycology, 2002, 14 (4): 281-292. |
| [15] | Bowker M A. Biological soil crust rehabilitation in theory and practice: An underexploited opportunity[J]. Restoration Ecology, 2007, 15 (1): 13-23. |
| [16] | Chen L Z, Xie Z M, Hu C X, et al. Man-made desert algal crusts as affected by environmental factors in Inner Mongolia, China[J]. Journal of Arid Environments, 2006, 67 (3): 521-527. |
| [17] | Xie Z M, Liu Y D, Hu C X, et al. Relationships between the biomass of algal crusts in fields and their compressive strength[J]. Soil Biology & Biochemistry, 2007, 39 (2): 567-572. |
| [18] | Liu Y D, Cockell C S, Wang G H, et al. Control of lunar and martian dust-experimental insights from artificial and natural cyanobacterial and algal crusts in the desert of Inner Mongolia, China[J]. Astrobiology, 2008, 8 (1): 75-86. |
| [19] | 饶本强, 吴易雯, 李华, 等. 库布齐沙漠不同发育类型人工结皮对露水凝结作用的比较研究[J]. 水土保持学报, 2011, 25 (6): 159-164. |
| [20] | 兰书斌, 胡春香, 饶本强, 等. 人工藻结皮形成过程中表土非降雨型水分吸收的变化情况[J]. 中国科学: 生命科学, 2010, 40 (8): 751-757. |
| [21] | 崔健宇, 江荣风. 土壤农化分析实验[M]. 北京: 中国农业大学出版社, 1998. |
| [22] | Wang W B, Liu Y D, Li D H, et al. Feasibility of cyanobacterial inoculation for biological soil crusts formation in desert area[J]. Soil Biology & Biochemistry, 2009, 41 (5): 926-929. |
| [23] | Schulten J A. Soil aggregation by cryptogams of a sand prairie[J]. American Journal of Botany, 1985, 72 (11): 1657-1661. |
| [24] | De Philippis R, Vincenzini M. Outermost polysaccharidic investments of cyanobacteria: nature, significance and possible applications[J]. Recent Research Development in Microbiology, 2003, 7: 13-22. |
| [25] | Lan S B, Wu L, Zhang D L, et al. Successional stages of biological soil crusts and their microstructure variability in Shapotou region (China)[J]. Environmental Earth Sciences, 2012, 65 (1): 77-88. |
| [26] | Büdel B, Darienko T, Deutschewith K, et al. Southern African biological soil crusts are ubiquitous and highly diverse in drylands, being restricted by rainfall frequency[J]. Microbial Ecology, 2009, 57 (2): 229-247. |
| [27] | Pickett S T A, McDonnell M J. Changing perspectives in community dynamics: a theory of successional forces[J]. Trends in Ecology & Evolution, 1989, 4 (8): 241-245. |
| [28] | Zaady E, Kuhn U, Wilske B, et al. Patterns of CO2 exchange in biological soil crusts of successional age[J]. Soil Biology & Biochemistry, 2000, 32 (7): 959-966. |