 2014, Vol.
2014, Vol. 



溶解硅酸盐作为一种主要的营养盐,支持了全球海洋中30%~40%的初级生产力[1],而在近岸和富营养水体更是支持了高达75%的初级生产力[2]. 生物硅的埋藏和溶解在Si的生物地球化学循环中起重要的作用,BSi在沉积物中的积累,可用于反映不同历史时期水体的富营养化及溶解态Si的消耗情况[3]. 生物硅的溶解是全球Si循环的重要环节,且Si循环与C循环关系密切. 为此,国际上开展了有关生物硅溶解动力学方面的研究. 并且生物硅溶解行为的研究主要集中在深海沉积物中,例如:大西洋[4,5,6]、 赤道太平洋[5]、 阿拉伯海等[7],特别是对南大洋沉积物中生物硅溶解行为进行了深入的研究[8,9,10,11]. 硅质细胞结构及其硅化程度、 温度、 pH、 盐度、 有机以及无机外壳、 水体硅酸盐的不饱和程度、 生物硅比表面积以及黏土矿物是影响生物硅溶解的主要因素[4,5,6,7,8,9,10,11,12]. 许多海洋生态系统已经证明了硅限制的存在,硅酸盐的再生对营养盐Si限制的海区是重要的补给,全球平均结果显示沉积物-水界面硅酸盐的再生量是河流输入量的10倍左右[13]. 硅酸盐的再生主要是通过生物硅的溶解,为了更好地了解Si的再生,因而有必要探究生物硅的溶解行为. 由于大洋水体较深以及水体中溶解硅酸盐浓度远远小于硅藻细胞骨架的溶解度[14],最终只有3%的生物硅埋藏在沉积物中[15, 16]; 而对于近岸水体,由于水深较浅,上层水体形成的生物硅能快速到达沉积物表层; 近岸沉积物中由于含有较多的矿物碎屑,其与生物硅的早期成岩作用致使铝硅酸盐自生矿物的沉积[17, 18]降低了生物硅表面反应活性,导致溶解度和溶解速率的降低[19, 20]. 同时生物扰动对于生物硅的溶解和保存有影响,会使溶解硅酸盐在沉积物-水界面通量增加[21, 22],也会将表层的新鲜硅藻转移到沉积物内部达20 cm[6],而使得生物硅保存下来.
长江作为世界第3大河,由于工农业排污和筑坝等因素,导致长江口溶解无机氮浓度相对于1960s增加 3倍[23],而活性硅酸盐浓度显著降低,增加了 Si限制的可能. 并导致在长江口及其邻近海域硅藻占整个浮游植物群落的比例由1984年的85%下降到2000年的60%,而从 2000~2005年每年有 30~80次的赤潮暴发[24]. 本研究通过批量培养实验(batch experiment)探讨长江口邻近海域沉积物中生物硅溶解行为.
2006年10月,随“北斗号”科学考察船对长江口邻近海域进行了调查,于16个站位(见图 1)采用箱式采泥器采集了柱状沉积物,并在现场对柱状沉积物进行分层,分层后的沉积物样一份冷冻保存,回到实验室后采用Christ®冷冻干燥机干燥,并称量干燥前后的重量; 另外一份用间隙水采样器Rhizon Sample采集间隙水样品,样品中加入饱和HgCl2,避光保存.
 | 图 1 柱状沉积物采样站位示意 Fig. 1 Sampling stations of core sediments |
生物硅批量培养实验用于测定生物硅的溶解速率常数. 称取一定量的沉积物放于盛有200 mL人工海水(0.7 mol ·L-1 NaCl,Tris+HCl pH=8.0)的50 mL塑料离心管中,振荡并定时取5 mL样品使用0.45 μm津腾®针头滤器过滤,保存到10 mL的带盖离心管中冷藏,样品用于测定溶液中的硅酸盐浓度以及溶解态Al的含量(因实验方法限制,只在最后时刻测定了溶解态Al浓度). 通过测定不同时刻硅酸盐浓度,可以获得硅酸盐浓度随时间的变化曲线(见图 2). 由图 2可知批量培养实验过程溶液中硅酸盐含量的增加与时间呈现明显的对数关系,生物硅在培养前期快速溶出,硅酸盐浓度增加较快,后期生物硅溶出较慢直至硅酸盐的浓度达到平衡.
 | 图 2 培养实验过程中,硅酸盐浓度随时间的变化曲线 Fig. 2 Changes of silicic acid concentrations during incubation periods |
沉积物中的生物硅采用改进的连续提取法:称取一定量的沉积物样品于50 mL的塑料离心管中. 在连续提取前使用HCl和H2 O2处理[25]沉积物样品以除去碳酸盐和有机相,加入去离子水离心清洗并干燥过夜,随后加入40 mL 2 mol ·L-1 Na2CO3,在85℃恒温水浴中连续提取8 h[26, 27],并于每小时离心取125 μL上清液,测定其中溶解硅酸盐浓度.
采用本研究的方法测定了太平洋深海沉积物样品,其生物硅含量为7.27%±0.057%(以SiO2计),方法的精密度为≤2.01%(n=5).
间隙水、 培养液以及提取液中的硅酸盐浓度采用硅钼蓝法[28]测定,提取液测定前先加入17.5 mL钼酸铵于比色管中消除干扰,方法的精密度为≤1.64%(n=7).
生物硅含量在不同深度沉积物中的空间分布如图 3所示,在0~0.5 cm、 0.5~1 cm、 1~1.5 cm、 1.5~2 cm沉积物中生物硅含量的平均值分别为:0.68%、 0.63%、 0.58%、 0.62%. 具有近岸含量高于远海的分布趋势,与宁修仁等[31]报道的黄、 东海初级生产力分布相似. Romero等[32]指出在秘鲁-智利上升流区域沉积物中生物硅和上层水体中初级生产力有比较好的正相关关系. 在长江口邻近海域、 长江口西北部以及浙闽沿岸海域生物硅含量较高,这与长江冲淡水向东海近岸输入大量的营养盐相关,同时也和沉积物类型、 沉积速率有关[33]. 位于近岸泥质区的站位,受大量陆源输入的影响,营养盐充足,初级生产力较高,沉积速率为1~2 cm ·a-1[34],且沉积物类型以黏土为主,颗粒物较细,细颗粒沉积物对有机物有着较大的富集能力[35],有机碳含量相对较高,有机碳含量高的沉积物有利于生物硅的保存[36]. DeMaster等[37]对罗斯海沉积物中生物硅的研究发现,生物硅在沉积物中的保存效率和沉积速率有比较好的正相关关系,沉积速率高的海域生物硅在水柱中的逗留时间较短,生物硅来不及溶解而被有效地埋藏到沉积中,沉积速率是影响生物硅在沉积物中保存的因素之一. 离岸较远的站位,初级生产力比较低,且沉积物类型为砂质沉积,沉积物颗粒较粗,其渗透性强,有机碳含量低,沉积速率较小,生物硅溶解较快,进而导致生物硅含量具有低值[38].
 | 图 3 不同深度沉积物中生物硅含量(SiO2%) 的平面分布 Fig. 3 Horizontal distribution of biogenic silica (SiO2%) in sediments of different depths |
长江口沉积物间隙水中硅酸盐随深度的分布如图 4所示,大多数站位间隙水中硅酸盐都维持在300 μmol ·L-1以下,除了H3-1与H3-16两个站位. H3-1站间隙水中硅酸盐由0.5 cm层的159.70 μmol ·L-1急剧增加至4.5 cm层处的492.80 μmol ·L-1,可能是由于0.5 cm层间隙水中硅酸盐相对于生物硅溶解度的不饱和程度较大为0.84[不饱和程度为间隙水中硅酸盐浓度和生物硅溶解度的差值与生物硅溶解度的比值. 合成硅在2℃海水中的溶解度(~900 μmol ·L-1 [39])或是被酸清除有机质外壳硅的残骸在4℃海水中的溶解度(~1 000 μmol ·L-1[40])被定义为生物硅的溶解度[7],本研究选择后者作为生物硅的溶解度]促进了生物硅的溶解,并随着生物硅溶解过程的进行,0.5 cm层以下间隙水中硅酸盐相对于生物硅溶解度的不饱和程度减小为3.5 cm层的0.51,这也和图 5中该站位生物硅的溶解速率常数随深度增加而递减相对应; 4.5 cm层以下硅酸盐呈现减小的趋势,该站位沉积物类型为砂质沉积并受到苏北沿岸流的影响,沉积环境不稳定,导致间隙水中硅酸盐变化比较剧烈. 对于间隙水中硅酸盐含量较高的H3-16站,各层次的硅酸盐浓度变化不大,大都在300~450 μmol ·L-1之间,该站位位于杭州湾的外海,生物硅含量较低,该站位属于砂质的沉积类型[41],黏土含量较少,这些因素有利于生物硅的溶解,致使间隙水中硅酸盐的浓度具有高值.
 | 图 4 间隙水中硅酸盐随深度的分布 Fig. 4 Vertical distribution of silicic acid in pore water |
 | 图 5 生物硅溶解速率常数随深度和站位变化 Fig. 5 Variations of BSi dissolution rate constant with stations and sediment depth |
间隙水中硅酸盐随深度的递增呈现增加的趋势,说明间隙水中硅酸盐未达到生物硅的溶解度,以生物硅的溶解作用为主,生物硅的溶解过程控制间隙水中硅酸盐具有这种分布趋势. 相反间隙水中硅酸盐随深度的递增呈现减小的趋势,可能是由于大量陆源碎屑的输入,陆源碎屑作为间隙水中Al离子的来源,为间隙水提供了丰富的Al离子,Al离子会吸附到生物硅矩阵的表面取替Si原子,减少生物硅表面的反应位点并与硅酸盐共沉淀形成硅铝酸盐,从而阻碍生物硅的溶解[11],此时共沉淀作用控制间隙水中硅酸盐浓度的变化. 同时本研究海域受到黄海沿岸流、 台湾暖流、 浙闽沿岸流以及长江冲淡水的影响,沉积环境不稳定,导致各站位间隙水中硅酸盐浓度变化比较剧烈. 不同站位间隙水中硅酸盐分布趋势变化也比较大,这可能和碎屑与生物硅含量比值的不同有关[12].
深海沉积物间隙水中的硅酸盐浓度随深度增加呈现增加的趋势,一般在10cm左右硅酸盐浓度达到一个渐近值[42],该深度以下硅酸盐浓度基本不变. 长江口邻近海域间隙水中硅酸盐浓度相对于南大洋、 赤道太平洋深海海域值偏小,随深度并不是都呈现典型增加的趋势,这可能和近岸沉积环境不稳定以及高的陆源碎屑输入相关,陆源碎屑与生物硅发生早期成岩作用阻碍生物硅的溶解[9,11,15],使得本研究区域间隙水中硅酸盐浓度较低.
Ehrenhauss等[43]对北海不同渗透性的沉积物间隙水中营养盐研究中发现,间隙水中硅酸盐浓度随沉积物类型渗透性的增强而递减,但是对于本研究海域并没有这种分布趋势,分析可能是由于间隙水中的Al或是陆架海水深较浅沉积环境不稳定造成本研究海域间隙水中硅酸盐具有随深度递增增大和减小的分布趋势.
Truesdale等[44]给出了批量培养实验中溶出的DSi随时间变化的关系式(用来解释批量培养实验中溶解速率常数的变化):
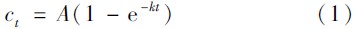
长江口邻近海域沉积物中生物硅溶解速率常数在0~0.5 cm、 0.5~1 cm、 1~1.5 cm、 1.5~2 cm沉积物中变化范围分别为: 0.58~3.90 d-1、 0.70~2.50 d-1、 1.36~2.90 d-1、 0.83~2.30 d-1; 随空间以及深度无明显规律(见图 5).
在H3-1和H3-4站位沉积物0~0.5 cm层生物硅的溶解速率常数具有高值,比其它站位高1~6倍,这可能是由于沉积物表层间隙水中硅酸盐浓度相对于生物硅溶解度的不饱和程度大都为0.84,从而加速了生物硅的溶解,也可能是由于这两个站位沉积物表层有新鲜硅藻的沉积,新鲜的硅藻具有高的比表面积和反应位点,在表层快速溶解. H3-1站生物硅的溶解速率常数随深度增加而递减,从该站间隙水中硅酸盐的剖面图(图 4)可以看出,前2 cm硅酸盐急剧增加,硅酸盐相对于生物硅溶解度的不饱和程度减小由0.84减为0.67,使得生物硅的溶解变慢.对于生物硅的溶解速率常数随深度增加有递增趋势的站位来说,分析可能是因为包裹在生物硅外面的有机外壳被浮游植物的摄食活动和微生物降解所清除[45],从而加速了生物硅的溶解,也可能是由于间隙水中Al随深度增加有减小的趋势,此时生物硅的溶解过程占主导. 通过图 6不同站位表层沉积物培养液不同时刻硅酸盐与最后培养液中溶解态Al浓度的变化关系可以看出, Al离子会影响生物硅的溶解行为,培养液中溶解态Al浓度较高的样品,其培养液中的硅酸盐浓度低. 说明在培养过程中铝会通过结合溶液中的溶解态硅形成铝硅酸盐降低硅酸盐的溶出速率,从而降低培养液中硅酸盐的浓度,这和H3-22站位具有高浓度的溶解态Al,但是其生物硅溶解速率常数具有低值相对应. Rickert[7]测定了不同Al浓度条件下生物硅的溶解速率,结果表明当溶解态Al浓度高时,溶解态Al和硅酸盐发生早期成岩作用,降低了生物硅表面活性,使得生物硅溶解生成的硅酸盐浓度和生物硅溶解速率低.
 | 图 6 各站位表层沉积物培养过程中不同时刻的硅酸盐浓度和最后时刻溶解态Al关系图 Fig. 6 Relationship between the temporal variations of silicic acid concentration and the final dissolved aluminum in surface sediments during the incubation period |
Van Cappellen 等[8, 9]在对南大洋沉积物中生物硅溶解动力学的研究中强调了间隙水中痕量元素Al对硅酸盐分布的影响,对于陆架边缘海而言,河流的输入是海水中Al的主要来源,其浓度相对于大洋海域较高,间隙水中Al浓度要远远高于上层水体中Al的浓度,闫丽等[46]对同航次的Al进行了研究,高值区主要分布在长江口以及舟山群岛附近,间隙水中Al主要来源于陆源碎屑的溶解,Al离子会吸附到生物硅的表面,阻碍其溶解,对本研究海域而言可能是由于间隙水中的Al浓度造成了生物硅溶解速率常数具有这种分布趋势,对其影响机制还需要进行深入的研究.
(1)长江口邻近海域沉积物中生物硅含量的水平分布呈现出近岸高于远海,南部海域高于北部海域的分布趋势,与初级生产力的分布趋势相似. 沉积物类型以及沉积速率会影响生物硅在沉积物中的保存.
(2)不同站位间隙水中硅酸盐的分布趋势并不相同,随深度并不是都呈现典型增加的趋势,且各站位硅酸盐浓度都在 500 μmol ·L-1以下,这可能和近岸沉积环境不稳定以及高的陆源碎屑输入相关,陆源碎屑与生物硅发生早期成岩作用阻碍生物硅的溶解,使得本研究区域间隙水中硅酸盐浓度较低.
(3)沉积物中生物硅溶解速率常数,随空间以及深度无明显规律,间隙水中硅酸盐相对于生物硅溶解度的不饱和程度以及痕量元素Al会影响本研究海域生物硅的溶解.
致谢: 感谢“北斗号”船员在调查期间给予的帮助,感谢中国海洋大学海洋生物地球化学实验室同学们的帮助,尤其曹璐在实验中给予的帮助.
| [1] | Buesseler K O. The decoupling of production and particulate export in the surface ocean[J]. Global Biogeochemical Cycles, 1998, 12 (2): 297-310. |
| [2] | Liu S M, Ye X W, Zhang J, et al. The silicon balance in Jiaozhou Bay, North China[J]. Journal of Marine Systems, 2008, 74 (1-2): 639-648. |
| [3] | Schelske C L, Conley D J, Stoermer E F. Biogenic silica and phosphorus accumulation in sediments as indices of eutrophication in the Laurentian Great Lakes[J]. Hydrobiologia, 1986, 143 (1): 79-86. |
| [4] | Ragueneau O, Gallinari M, Corrin L, et al. The benthic silica cycle in the Northeast Atlantic: annual mass balance, seasonality, and importance of non-steady-state processes for the early diagenesis of biogenic opal in deep-sea sediments[J]. Progress in Oceanography, 2001, 50 (1-4): 171-200. |
| [5] | Gallinari M, Ragueneau O, Corrin L, et al. The importance of water column processes on the dissolution properties of biogenic silica in deep-sea sediments I. Solubility[J]. Geochimica et Cosmochimica Acta, 2002, 66 (15): 2701-2717. |
| [6] | Gallinari M, Ragueneau O, DeMaster D J, et al. Influence of seasonal phytodetritus deposition on biogenic silica dissolution in marine sediments-Potential effects on preservation[J]. Deep Sea Research Part Ⅱ: Topical Studies in Oceanography, 2008, 55 (22-23): 2451-2464. |
| [7] | Rickert D. Dissolution kinetics of biogenic silica in marine environments[D]. Germany: Alfred Wegener Institute for Polar and Marine Research, 2000. 1-234. |
| [8] | Van Cappellen P, Qiu L. Biogenic silica dissolution in sediments of the Southern Ocean. Ⅰ. Solubility[J]. Deep Sea Research Part Ⅱ: Topical Studies in Oceanography, 1997, 44 (5): 1109-1128. |
| [9] | Van Cappellen P, Qiu L. Biogenic silica dissolution in sediments of the Southern Ocean. Ⅱ. Kinetics[J]. Deep Sea Research Part Ⅱ: Topical Studies in Oceanography, 1997, 44 (5): 1129-1149. |
| [10] | Van Cappellen P. Reactive surface area control of the dissolution kinetics of biogenic silica in deep-sea sediments[J]. Chemical Geology, 1996, 132 (1-4): 125-130. |
| [11] | Dixit S, Van Cappellen P, Van Bennekom A J. Processes controlling solubility of biogenic silica and pore water build-up of silicic acid and pore water build-up of silicic acid in marine sediments[J]. Marine Chemistry, 2001, 73 (3-4): 333-352. |
| [12] | Rickert D, Schlüter M, Wallmann K. Dissolution kinetics of biogenic silica from the water column to the sediments[J]. Geochimica et Cosmochimica Acta, 2002, 66 (3): 439-455. |
| [13] | Tréguer P J, De La Rocha C L. The world ocean silica cycle[J]. Annual Review of Marine Science, 2013, 5 (1): 477-501. |
| [14] | Hurd D C. Interactions of biogenic opal, sediment and seawater in the Central Equatorial Pacific[J]. Geochimica et Cosmochimica Acta, 1973, 37 (10): 2257-2266. |
| [15] | Nelson D M, Tréguer P, Brzezinski M A, et al. Production and dissolution of biogenic silica in the ocean: Revised global estimates, comparison with regional data and relationship to biogenic sedimentation[J]. Global Biogeochemical Cycles, 1995, 9 (3): 359-372. |
| [16] | Tréguer P, Nelson D M, Van Bennekom A J, et al. The silica balance in the world ocean: a reestimate[J]. Science, 1995, 268 (5209): 375-379. |
| [17] | Michalopoulos P, Aller R C, Reeder R J. Conversion of diatoms to clays during early diagenesis in tropical, continental shelf muds[J]. Geology, 2000, 28 (12): 1095-1098. |
| [18] | Michalopoulos P, Aller R C. Early diagenesis of biogenic silica in the Amazon delta: Alteration, authigenic clay formation, and storage[J]. Geochimica et Cosmochimica Acta, 2004, 68 (5): 1061-1085. |
| [19] | Van Bennekom A J, Fred Jansen J H, Van der Gaast S J, et al. Aluminium-rich opal: an intermediate in the preservation of biogenic silica in the Zaire (Congo) deep-sea fan[J]. Deep Sea Research Part A. Oceanographic Research Papers, 1989, 36 (2): 173-190. |
| [20] | Van Bennekom A J, Buma A G J, Nolting R F. Dissolved aluminium in the Weddell-Scotia Confluence and effect of Al on the dissolution kinetics of biogenic silica[J]. Marine Chemistry, 1991, 35 (1-4): 423-434. |
| [21] | Ragueneau O, Chsuvaud L, Leynaert A, et al. Direct evidence of a biologically active coastal silicate pump: Ecological implications[J]. Limnology and Oceanography, 2002, 47 (6): 1849-1854. |
| [22] | Ragueneau O, Chauvaud L, Leynaert A, et al. Biodeposition by an invasive suspension feeder impacts the biogeochemical cycle of Si in a coastal ecosystem (Bay of Brest, France)[J]. Biogeochemistry, 2005, 75 (1): 19-41. |
| [23] | Li M T, Xu K Q, Watanabe M, et al. Long-term variations in dissolved silicate, nitrogen, and phosphorus flux from the Yangtze River into the East China Sea, and impacts on estuarine ecosystem[J]. Estuarine, Coastal and Shelf Science, 2007, 71 (1-2): 3-12. |
| [24] | Zhou M J, Shen Z L, Yu R C. Responses of a coastal phytoplankton community to increased nutrient input from the Changjiang (Yangtze) River[J]. Continental Shelf Research, 2008, 28 (12): 1483-1489. |
| [25] | Mortlock R A, Froelich P N. A simple method for the rapid determination of biogenic opal in pelagic marine sediments[J]. Deep Sea Research Part A. Oceanographic Research Papers, 1989, 36 (9): 1415-1426. |
| [26] | Liu S M, Ye X W, Zhang J, et al. Problems with biogenic silica measurement in marginal seas[J]. Marine Geology, 2002, 192 (4): 383-392. |
| [27] | Liu S M, Zhang J, Li R X. Ecological significance of biogenic silica in the East China Sea[J]. Marine Ecology Process Series, 2005, 290: 15-26. |
| [28] | Strickland J D H, Parsons T R. Determination of reactive nitrite. In: A Practical Handbook of Seawater Analysis[M]. Ottawa, Fisheries Research Board of Canada, 1968. 71-75. |
| [29] | Ren J L, Zhang J, Luo J Q, et al. Improved fluorimetric determination of dissolved aluminium by micelle-enhanced lumogallion complex in natural waters[J]. Analyst, 2001, 126 (5): 698-702. |
| [30] | Hydes D J, Liss P S. Fluormetric method for the determination of low concentrations of dissolved aluminium in natural waters[J]. Analyst, 1976, 101: 922-931. |
| [31] | 宁修仁, 刘子琳, 蔡昱明. 我国海洋初级生产力研究二十年[J]. 海洋学研究, 2000, 18 (3): 13-20. |
| [32] | Romero O, Hebbeln D. Biogenic silica and diatom thanatocoenosis in surface sediments below the Peru-Chile Current: controlling mechanisms and relationship with productivity of surface waters[J]. Marine Micropaleontology, 2003, 48 (1-2): 71-90. |
| [33] | Bernárdex P, Francés G, Prego R. Benthic-pelagic coupling and postdepositional processes as revealed by the distribution of opal in sediments: The case of the Ría de Vigo (NW Iberian Peninsula)[J]. Estuarine, Coastal and Shelf Science, 2006, 68 (1-2): 271-281. |
| [34] | Liu J P, Li A C, Xu K H, et al. Sedimentary features of the Yangtze River-derived along-shelf clinoform deposit in the East China Sea[J]. Continental Shelf Research, 2006, 26 (17-18): 2141-2156. |
| [35] | 王华新, 线薇微. 长江口表层沉积物有机碳分布及其影响因素[J]. 海洋科学, 2011, 35 (5): 24-31. |
| [36] | DeMastr D J. The accumulation and cycling of biogenic silica in the Southern Ocean: revisiting the marine silica budget[J]. Deep Sea Research Part Ⅱ, Topical Studies in Oceanography, 2002, 49 (16): 3155-3167. |
| [37] | DeMaster D J, Ragueneau O, Nittrouer C A. Preservation efficiencies and accumulation rates for biogenic silica and organic C, N, and P in high-latitude sediments: The Ross Sea[J]. Journal of Geophysical Research: Oceans, 1996, 101 (C8): 18501-18518. |
| [38] | 李学刚, 宋金明, 袁华茂, 等. 胶州湾沉积物中高生源硅含量的发现——胶州湾浮游植物生长硅限制的证据[J]. 海洋与湖沼, 2005, 36 (6): 572-579. |
| [39] | Willey J D. Effects of aging on silica solubility: a laboratory study[J]. Geochimica et Cosmochimica Acta, 1980, 44 (4): 537-578. |
| [40] | Hurd D C. Interactions of biogenic silica, sediment and seawater in the Central Equatorial Pacific[J]. Geochimica et Cosmochimica Acta, 1973, 37 (10): 2267-2282. |
| [41] | 于培松, 薛斌, 潘建明, 等. 长江口和东海海域沉积物粒径对有机质分布的影响[J]. 海洋学研究, 2011, 29 (3): 202-208. |
| [42] | Rabouille C, Gaillard J, Tréguer P, et al. Biogenic silica recycling in surficial sediments across the Polar Front of the Southern Ocean (Indian Sector)[J]. Deep-Sea Research Part Ⅱ: Topical Studies in Oceanography, 1997, 44 (5): 1151-1176. |
| [43] | Ehrenhauss S, Witte U, Bühring S L, et al. Effect of advective pore water transport on distribution and degradation of diatoms in permeable North Sea sediments[J]. Marine Ecology Progress Series, 2004, 271: 99-111. |
| [44] | Truesdale V W, Greenwood J E, Rendell A. The rate-equation for biogenic silica dissolution in seawater-new hypotheses[J]. Aquatic Geochemistry, 2005, 11 (3): 319-343. |
| [45] | Patrick S, Holding A J. The effect of bacteria on the solubilization of silica in diatom frustules[J]. Journal of Applied Bacteriology, 1985, 59 (1): 7-16. |
| [46] | 闫丽, 任景玲, 罗建, 等. 长江口邻近海域溶解态铝的分布及季节变化[J]. 中国海洋大学学报(自然科学版), 2011, 41 (增刊): 259-266. |