 2014, Vol.
2014, Vol. 



2. 广东省生态环境与土壤研究所, 广州 510650;
3. 湖南农业大学教育学院, 长沙 410128
2. Guangdong Institute of Eco-Environmental and Soil Sciences, Guangzhou 510650, China;
3. College of Education, Hunan Agricultural University, Changsha 410128, China
甲烷(CH4)对全球变暖的贡献率高达20%,其单分子增温效应是二氧化碳(CO2)的20倍,是仅次于CO2的重要温室气体[1]. 稻田是主要的CH4排放源,年均排放量约为20~100 Tg[2]. 稻田CH4释放受到多种因素的影响,如稻田水分管理(干湿交替)、 土壤电子受体[Fe(Ⅲ)等]等对CH4排放产生抑制作用[3]. 这种抑制作用使水分管理和添加外源电子受体等方式成为减缓稻田CH4排放的潜在途径,且其作用机制日益受到重视.
铁作为土壤中极为重要的变价元素[4,5,6],存在价态主要有Fe(Ⅱ)与Fe(Ⅲ),在自然环境中的存在形态多种多样,如水铁矿、 赤铁矿、 菱铁矿、 硫铁矿等[4]. 土壤中存在着大量铁呼吸菌,它们能够利用Fe(Ⅲ)作为末端电子受体氧化有机质,进行铁呼吸获得能量从而完成生命活动[7, 8]. Lovley[9]在1991年发现,铁呼吸菌相对于产甲烷菌更具底物竞争优势,铁呼吸过程可以抑制CH4产生. 大量研究证实厌氧水稻土中Fe(Ⅲ)呼吸是抑制CH4产生的重要原因[10,11,12]. 铁矿物的晶型直接影响Fe(Ⅲ)的还原活性和生物可利用性,晶型较低的铁矿物更容易被铁呼吸菌利用. 前期研究表明晶型较低的水铁矿比晶型较高的纤铁矿、 赤铁矿等更易被微生物利用还原,且对产CH4的抑制作用更为显著[13].
稻田特殊的管理方式是淹水排水频繁交替(即干湿交替),干湿交替的排水过程中O2的输入可加速水稻土有机碳的有氧矿化[14,15,16]. Ma等[3]和Wassmann等[17]研究表明干湿交替能显著抑制产甲烷菌活性,从而减少水稻土CH4的生成和排放. 同理,干湿交替的排水过程中进入土壤的O2也能够将淹水过程中形成的Fe(Ⅱ)通过化学氧化作用再生成Fe(Ⅲ),从而使淹水阶段Fe(Ⅲ)呼吸的持续进行成为可能[18, 19]. 但是在干湿交替这种典型的稻田水分管理模式下,关于不同晶型铁氧化物对水稻土CH4排放的影响还鲜见报道.
本实验采用室内恒温培养试验,经过连续3次淹水-排水处理,研究了干湿交替条件下两种不同晶型铁氧化物(水铁矿和赤铁矿)对华南水稻土CH4排放的影响,以期进一步探明水稻田CH4排放与田间水分管理及土壤铁氧化物之间的相互关系.
供试土壤取自华南水稻田(23°10.099′N,113°21.696′E),自然风干后,捡去土中水稻根及异物,碾碎过10目筛,充分混匀,经分析土壤质地为黏质壤土. 土壤基本理化性质见表 1.
| 表 1 供试土样的基本理化性质 Table 1 Physical and chemical characteristics of the tested soil |
试验设计共6个处理,分别为:①CK-CF(土壤,持续淹水); ②SF-CF(土壤+水铁矿,持续淹水); ③SH-CF(土壤+赤铁矿,持续淹水); ④CK-DW(土壤,干湿交替); ⑤SF-DW(土壤+水铁矿,干湿交替); ⑥SH-DW(土壤+赤铁矿,干湿交替). 每个处理有3个重复. 每个处理的土壤干重为20 g,以40 mL去离子水与土壤按水土比2 ∶1混合于125 mL培养瓶中. 水铁矿、 赤铁矿的添加量均为每克干土40 μmol. 干湿交替的处理为:淹水期8~10 d,排水期6 d,再次淹水,以此重复3个周期,25℃下恒温避光培养46 d. 分别在培养第4、 11、 17、 19、 25、 31、 40和46 d破坏性取样. 为保证产CH4过程的持续进行,在第一个干湿交替周期结束后分别向各处理加入15 mmol ·L-1乙酸.
试验中所用的铁氧化物合成参照文献[20].
气体采样次数参照文献[3]. 每个淹水排水期气体采样的具体时间为:各处理在第一个淹水期均隔天采样,即整个培养期第1、 3、 5、 7、 9、 11 d采集气样; 第一个排水期采样3次,即整个培养期第13、 15、 17 d采样; 第二个淹水期采样3次,即整个培养期19、 21、 25 d采样; 第二个排水期采样2次,即整个培养期第27、 31 d采样; 第三个淹水期及第三个排水期均采样1次,即整个培养期第40 d和第46 d采样. 采样时,将培养瓶密封培养2 h,用气密性注射器分别抽取t1=0和t2=2 h时培养瓶内的气体200 μL,两者之差即为2 h内的气样通量. 通过带有氢离子火焰检测器的气相色谱仪器(GC7900)进行气体分析(进样口温度:120℃,柱温:80℃,FID检测器:350℃). CH4通量[FCH4,μmol ·(g ·h)-1]的计算[21]见公式(1).
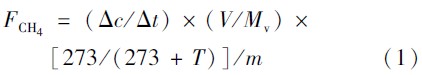
产CH4潜力检测:破坏性取样1 h内,取10 g鲜土于125 mL培养瓶中,补充10 mL除氧无菌水,100% N2冲洗培养瓶20 min,于25℃暗处培养10 d,每2 d取样测气. 产CH4潜力计算方法参照文献[3].
Fe(Ⅱ)和Fe(Ⅲ)测定[22]:用一次性无菌注射器吸取2 mL水土混合样,加入8 mL 0.5 mol ·L-1盐酸溶液,振荡1.5 h后加入4 mL 0.1%乙酸钠溶液和4 mL 0.1%邻菲罗啉溶液中显色,4 000 r ·min-1离心后采用分光光度法测定Fe(Ⅱ)浓度. 用一次性无菌注射器吸取2 mL水土混合样,加入2 mL 10%盐酸羟胺,再加入4 mL 10%乙酸钠溶液和4 mL 0.1%邻菲罗啉显色, 4 000 r ·min-1离心后采用分光光度法测定总Fe浓度. Fe(Ⅲ)浓度即为总Fe浓度和Fe(Ⅱ)浓度之差.
采用 SPSS 16.0进行ANOVA分析,检验各检测指标在各处理间的显著性差异; 对不同水分和铁氧化物处理下的CH4排放速率、 产CH4潜力进行多因素方差分析,并对CH4排放速率、 产CH4潜力与Fe(Ⅱ)及Fe(Ⅲ)含量之间进行相关性分析.
由图 1所示,除SF外,所有处理的CH4排放速率在培养第6 d或第8 d后达到峰值[约20~25 μmol ·(g ·h)-1],在培养第17 d后即第一个干湿交替周期结束时趋于零,为保证产CH4过程的持续进行,于培养第17 d向各处理中添加浓度为15 mmol ·L-1的乙酸,因此所有处理在第二个干湿交替周期达到的最大CH4排放速率都明显高于第一个周期. 培养46 d后,所有处理的CH4排放速率降低至零.
 | 图 1 CF及DW条件下添加不同铁氧化物后CH4排放速率的动态变化 Fig. 1 Rates of methane emission after adding different iron oxides during CF and DW |
由图 1可知,与CF相比,DW对水稻土CH4排放速率有十分显著的影响. DW的CH4平均排放速率分别比CK降低了61%. 通过对比添加不同铁氧化物对CH4排放速率的影响可知,整个培养期内SF的CH4排放速率最低,与CK相比,平均降低了83%. SH的CH4排放速率随时间的变化规律与CK一致,但其均值比CK-CF低约22%(图 1).
将DW与铁氧化物的耦合对CH4排放速率的抑制作用进行方差分析可知,DW与铁氧化物对CH4排放速率的耦合抑制非常显著(R2=0.93,P<0.01,表 2). 在整个培养阶段,以CK-CF为参照,铁氧化物与DW的耦合抑制了CH4平均排放速率达80%,高于DW与铁氧化物二者单独对CH4平均排放速率的抑制作用(61%和53%),其中,SH-DW对CH4平均排放速率的抑制程度为65%,而SF-DW则抑制了94%的CH4排放速率,即干湿交替耦合晶型较弱的水铁矿对CH4排放的抑制效果比干湿交替耦合晶型较好的赤铁矿更加突出.
| 表 2 不同条件下CH4排放速率方差模型分析结果 Table 2 Between-subject effects of methane emission rates under different conditions |
由图 2可知,各处理的产CH4潜力随时间的变化趋势与CH4排放速率大致相同. 除SF外,所有处理的产CH4潜力在培养第11 d达到峰值[约350 μmol ·(g ·h)-1]. 于培养第17 d添加乙酸后,包括SF的所有处理的产CH4潜力在培养第20 d或第25 d达到峰值. 培养后期,所有处理的产CH4潜力处于持续降低趋势.
 | 图 2 CF及DW条件下添加不同铁氧化物后产CH4潜力的动态变化 Fig. 2 Methane production potential after adding different iron oxides during CF and DW |
对比图 2可知,DW对水稻土产CH4潜力有明显的抑制作用. 整个培养期内,DW的产CH4潜力均值比CF下降了53%. 对比不同铁氧化物对水稻土产CH4潜力可知,SF的产CH4潜力在整个培养期内显著低于CK,其产CH4潜力均值比CK降低了86%,而SH的产CH4潜力随时间的变化规律与CK的一致,其均值比CK降低了6%.
由表 3可知,DW与铁氧化物对产CH4潜力的耦合效应极显著(R2=0.88,P<0.01). 与CK-CF相比,整个培养期内DW与铁氧化物耦合对产CH4潜力的抑制效果达75%,均高于DW或铁氧化物单独对产CH4潜力的抑制效果(53%和46%). 其中,SH-DW的产CH4潜力均值降低了57%,而SF-DW对水稻土产CH4潜力降低程度达到了93%. 由此可知,干湿交替耦合晶型较弱的水铁矿比耦合晶型较好的赤铁矿对水稻土产CH4潜力有更为明显的抑制作用.
| 表 3 不同条件下产CH4潜力方差模型分析结果 Table 3 Between-subject effects of methane production potential under different conditions |
由图 3可知,在不同水分处理下,SH的Fe(Ⅱ)含量变化趋势均与CK接近,且随培养时间的波动幅度较小. 整个培养期内, SF的Fe(Ⅱ)含量总体呈上升趋势,且SF的Fe(Ⅱ)含量显著高于SH和CK.
 | 图 3 CF及DW条件下添加不同铁氧化物后Fe(Ⅱ)浓度的动态变化 Fig. 3 Changes of Fe(Ⅱ) concentration after adding different iron oxides during CF and DW |
纵观整个培养期,CK-DW、 SH-DW、 SF-DW中的Fe(Ⅱ)含量随时间波动幅度明显大于各CF组(图 3). CK-DW和SH-DW的Fe(Ⅱ)含量呈现较小幅度的周期性波动,而SF-DW的Fe(Ⅱ)含量随着排水淹水的交替进行呈现出极为显著的周期性波动,且总体呈缓慢上升趋势[图 3(b)]. CK-CF和SH-CF的Fe(Ⅱ)含量在培养期46 d内都维持在0.7 mmol ·g-1左右,SF-CF的Fe(Ⅱ)含量呈平缓上升趋势[图 3(a)]. 由表 4可知,不同处理间的Fe(Ⅱ)含量与CH4排放速率及产CH4潜力呈显著负相关.
| 表 4 CF与DW条件下铁元素价态与CH4排放速率、产CH4潜力的相关关系 1) Table 4 Correlations between iron oxides and methane emission rates, methane production potential under DW/CF conditions |
如图 4所示,在不同水分处理下,CK的Fe(Ⅲ)含量最低,且在培养期46 d内一直处于平稳状态(约0.06 mmol ·g-1). 尽管SH的Fe(Ⅲ)含量较高,但在培养期内并无明显波动(约0.6 mmol ·g-1). SF的Fe(Ⅲ)含量的初始值最高,约1.0 mmol ·g-1,但从培养开始,SF的Fe(Ⅲ)含量一直处于下降趋势,培养46 d后降至0.2~0.4 mmol ·g-1.
 | 图 4 CF及DW条件下添加不同铁氧化物后Fe(Ⅲ)浓度的动态变化 Fig. 4 Changes of Fe(Ⅲ) concentration after adding different iron oxides during CF and DW |
对比图 4可知,CK-DW和SH-DW的Fe(Ⅲ)含量与持续淹水处理接近,且变化趋势一致. SF-DW的Fe(Ⅲ)含量波动幅度则明显强于持续淹水, 其Fe(Ⅲ)含量则随着淹水排水的交替进行呈现周期性的波动,且随着培养时间的推移呈下降趋势,培养46 d后降至0.4 mmol ·g-1. SF-CF的Fe(Ⅲ)含量呈平缓下降趋势,培养46 d后降至0.2 mmol ·g-1. 由表 4可知,Fe(Ⅲ)含量与CH4排放速率及产CH4潜力呈显著负相关.
从实验结果可以看出,干湿交替耦合不同铁氧化物对CH4排放的抑制作用超过了干湿交替或铁氧化物单独对CH4排放的抑制作用. 这种差异可能是因为干湿交替的水分管理模式驱动了水稻土中Fe(Ⅱ)/Fe(Ⅲ)的循环,使得铁呼吸对水稻土产CH4过程形成多次抑制效应. 铁呼吸早已被确认是微生物以胞外固态铁氧化物作为末端电子受体,通过氧化电子供体偶联Fe(Ⅲ)还原,并获得能量的厌氧呼吸形式[22]. 大量研究表明,铁呼吸菌相对于产甲烷菌对产甲烷前体(乙酸、 H2等)更具有竞争优势,因此铁呼吸过程可以抑制CH4的产生[9,13,24]. 前期关于干湿交替对CH4影响的研究多集中于O2的直接影响,认为干湿交替的排水过程导致土壤中的O2含量增加破坏了产甲烷菌赖以生存的厌氧环境[25],并促进甲烷氧化菌有氧代谢[26,27,28],从而抑制了CH4排放,但都未涉及Fe(Ⅲ)/Fe(Ⅱ)循环在干湿交替过程中对CH4排放的影响. 大量研究表明,频繁的降水会导致土壤氧化-还原状态交替发生,强化Fe(Ⅲ)的循环再生,提高了铁呼吸强度[32,30,31,27]. 与频繁降水的效果类似,稻田干湿交替同样导致土壤氧化-还原状态的交替发生. 淹水期的厌氧环境下,铁氧化物在铁呼吸菌的作用下被还原成Fe(Ⅱ),存留于土壤中的Fe(Ⅱ)在排水期被扩散进土壤中的O2迅速氧化成Fe(Ⅲ)[33],Fe(Ⅲ)在下一个淹水期作为铁呼吸的电子受体继续消耗土壤中的H2及乙酸盐等产甲烷基质. 因此,干湿交替可通过化学氧化还原作用加速Fe(Ⅲ)的循环再生,促使铁呼吸在多个淹水期的持续进行,进而抑制CH4排放.
值得关注的是,培养期内干湿交替与晶型较弱的水铁矿耦合对CH4排放速率的抑制效果明显高于干湿交替与晶型较高的赤铁矿的耦合效应. 从氧化还原电位方面来看,赤铁矿的氧化还原电位(Eh)约为 -287 mV,而水铁矿的Eh为-100~100 mV[33],而产甲烷菌开始大量产生CH4的Eh值低于-150 mV[16, 17],因此水铁矿提高了土壤的Eh值,对CH4生成有抑制效应[13]. 从2.3节可知,干湿交替3个处理中,水铁矿组的Fe(Ⅱ)含量和周期性波动幅度远高于空白对照组和赤铁矿组,这说明土壤中的水铁矿还原比赤铁矿还原更为剧烈,从而抑制产CH4潜力和CH4排放速率的效果更为显著. 原因可能在于铁呼吸菌对氧化铁的还原具有一定选择性. 曲东等[35]在研究不同氧化铁的微生物还原时发现,微生物通常更易利用晶型较弱的水铁矿,而对晶型较高的赤铁矿的还原性不强. 此外,Weiss等[19]在研究热带森林土壤的微生物铁还原潜力时也发现,晶型较弱的铁氧化物更易被铁呼吸菌利用. 因此,本实验中干湿交替条件下,水铁矿组的铁呼吸强度远大于赤铁矿组,导致水铁矿组的产CH4潜力最低,其CH4排放速率也最低.
干湿交替强化水稻土Fe(Ⅲ)呼吸抑制CH4排放的作用机制如图 5所示:在干湿交替水分管理模式下,排水期进入土壤的O2氧化Fe(Ⅱ)生成Fe(Ⅲ),这些再生的Fe(Ⅲ)在下一个淹水期被铁呼吸菌作为电子受体利用,与产甲烷菌竞争乙酸和H2等产甲烷前体,从而抑制产CH4过程. 从2.3节可以看出,干湿交替下,水铁矿和赤铁矿处理的Fe(Ⅲ)/Fe(Ⅱ)含量随着淹水-排水的交替进行分别呈现出明显的升高-降低和下降-上升的周期性波动,且SF的Fe(Ⅱ)和Fe(Ⅲ)变化幅度明显大于SH. 这也就说明干湿交替确实可以促进Fe(Ⅲ)/Fe(Ⅱ)循环,Fe(Ⅲ)的循环再生保证了铁呼吸在排水后的淹水期的持续进行,进而更大程度地抑制产CH4潜力和CH4排放速率,并且这种抑制效果在与晶型较弱的水铁矿耦合中显得尤为突出.
 | 图 5 干湿交替强化水稻土Fe(Ⅲ)呼吸对CH4排放的抑制作用机制 Fig. 5 Proposed mechanism for suppression of methanogenesis by enhanced Fe(Ⅲ) respiration under DW in paddy soil |
(1)由于干湿交替促进了Fe(Ⅲ)/Fe(Ⅱ)的循环,保证铁呼吸在多个淹水期的持续进行,促使铁呼吸菌不断消耗土壤中H2及乙酸盐等产甲烷基质,从而干湿交替耦合铁氧化物对稻田CH4排放的抑制作用超过了干湿交替或铁氧化物单独对CH4排放的抑制作用.
(2)干湿交替与铁氧化物耦合过程中,由于水铁矿的Eh值明显高于产CH4时所需Eh值,且晶型较弱的水铁矿比晶型较高的赤铁矿更易被微生物还原,因此干湿交替与水铁矿耦合对稻田CH4排放的抑制效果明显高于干湿交替与赤铁矿的耦合效应.
| [1] | WMO. Greenhouse Gas Bulletin: The State of Greenhouse Gases in the Atmosphere based on Global Observations through 2011[EB/OL]. 2012-11-19. |
| [2] | Cai Z C, Xing G X, Yan X Y, et al. Methane and nitrous oxide emissions from rice paddy fields as affected by nitrogen fertilisers and water management[J]. Plant and Soil, 1997, 196 (1): 7-14. |
| [3] | Ma K, Conrad R, Lu Y H. Responses of methanogen mcrA genes and their transcripts to an alternate dry/wet cycle of paddy field soil[J]. Applied and Environmental Microbiology, 2012, 78 (2): 445-454. |
| [4] | Kappler A, Straub K L. Geomicrobiological cycling of iron[J]. Reviews in Mineralogy and Geochemistry, 2005, 59 (1): 85-108. |
| [5] | Straub K L, Schönhube W A, Buchholz-Cleven B E E, et al. Diversity of ferrous iron-oxidizing, nitrate-reducing bacteria and their involvement in oxygen-independent iron cycling[J]. Geomicrobiology Journal, 2004, 21 (6): 371-378. |
| [6] | Wang J J, Muyzer G, Bodelier P L E, et al. Diversity of iron oxidizers in wetland soils revealed by novel 16S rRNA primers targeting Gallionella-related bacteria[J]. International Society for Microbial Ecology, 2009, 3 (6): 715-725. |
| [7] | Kim D, Strathmann T J. Role of organically complexed iron(Ⅱ) species in the reductive transformation of RDX in anoxic environments[J]. Environmental Science and Technology, 2007, 41 (4): 1257-1264. |
| [8] | Weber K A, Achenbach L A, Coates J D. Microorganisms pumping iron: anaerobic microbial iron oxidation and reduction[J]. Nature Reviews Microbiology, 2006, 4 (10): 752-764. |
| [9] | Lovley D R. Dissimilatory Fe(Ⅲ) and Mn(Ⅳ) reduction[J]. Microbiology Review, 1991, 55 (2): 259-287. |
| [10] | Conrad R. Contribution of hydrogen to methane production and control of hydrogen concentrations in methanogenic soils and sediments[J]. FEMS Microbiology Ecology, 1999, 28 (3): 193-202. |
| [11] | Yao H, Conrad R. Thermodynamics of methane production in different rice paddy soils from China, the Philippines and Italy[J]. Soil Biology and Biochemistry, 1999, 31 (3): 463-473. |
| [12] | Achtnich C, Bak F, Conrad R. Competition for electron-donors among nitrate reducers, ferric iron reducers, sulfate reducers, and methanogens in anoxic paddy soil[J]. Biology and Fertility of Soils, 1995, 19 (1): 65-72. |
| [13] | 曲东, Schnell S. 外源氧化铁对水稻土甲烷形成的抑制[J]. 环境科学学报, 2002, 22 (1): 65-69. |
| [14] | 刘艳丽, 张斌, 胡锋, 等. 干湿交替对水稻土碳氮矿化的影响[J]. 土壤, 2008, 40 (4): 554-560. |
| [15] | Mikha M M, Rice C W, Milliken G A. Carbon and nitrogen mineralization as affected by drying and wetting cycles[J]. Soil Biology and Biochemistry, 2005, 37 (2): 339-347. |
| [16] | Harrison-Kirk T, Beare M H, Meenken E D, et al. Soil organic matter and texture affect responses to dry/wet cycles: Effects on carbon dioxide and nitrous oxide emissions[J]. Soil Biology and Biochemistry, 2013, 57: 43-55. |
| [17] | Wassmann R, Lantin R S, Neue H U, et al. Characterization of methane emissions from rice fields in Asia. Ⅲ. Mitigation options and future research needs[J]. Nutrient Cycling in Agroecosystems, 2000, 58 (1-3): 23-36. |
| [18] | Fimmen R L, deB Vasudevan D Jr, Williams M A, et al. Rhizogenic Fe-C redox cycling: a hypothetical biogeochemical mechanism that drives crustal weathering in upland soils[J]. Biogeochemistry, 2008, 87 (2): 127-141. |
| [19] | Weiss J V, Emerson D, Backer S M, et al. Enumeration of Fe(Ⅱ)-oxidizing and Fe(Ⅲ)-reducing bacteria in the root zone of wetland plants: Implications for a rhizosphere iron cycle[J]. Biogeochemistry, 2003, 64 (1): 77-96. |
| [20] | Schwertmann U, Cornell R M. Iron oxides in the laboratory: preparation and characterization[M]. Wernheim: VCH, 1991.61-109. |
| [21] | Rolston D E. Gas flux[A]. In: Klute A (ed.). Methods of Soil Analysis, part 1, (2nd ed.)[C]. Agron. Monogr. 9. Madison: American Society of Agronomy and Soil Science Society of America, WI, 1986. 1103-1119. |
| [22] | Roden E E, Zachara J M. Microbial reduction of crystalline iron(Ⅲ) oxides: Influence of oxide surface area and potential for cell growth[J]. Environmental Science and Technology, 1996, 30 (5): 1618-1628. |
| [23] | Lovley D R. Organic matter mineralization with the reduction of ferric iron: a review[J]. Geomicrobiology Journal, 1987, 5 (3-4): 375-399. |
| [24] | Jäckel U, Schnell S. Suppression of methane emission from rice paddies by ferric iron fertilization[J]. Soil Biology and Biochemistry, 2000, 32 (11-12): 1811-1814. |
| [25] | 张军科, 江长胜, 郝庆菊, 等. 耕作方式对紫色水稻土农田生态系统CH4和N2O排放的影响[J]. 环境科学, 2012, 33 (6): 1978-1986. |
| [26] | Denef K, Six J, Bossuyt H, et al. Influence of dry-wet cycles on the interrelationship between aggregate, particulate organic matter, and microbial community dynamics[J]. Soil Biology and Biochemistry, 2001, 33 (12): 1599-1611. |
| [27] | Appel T. Non-biomass soil organic N-the substrate for N mineralization flushes following soil drying-rewetting and for organic N rendered CaCl2-extractable upon soil drying[J]. Soil Biology and Biochemistry, 1998, 30 (10-11): 1445-1456. |
| [28] | Van Gestel M, Merckx R, Vlassak K. Microbial biomass responses to soil drying and rewetting: the fate of fast-and slow-growing microorganisms in soils from different climates[J]. Soil Biology and Biochemistry, 1993, 25 (1): 109-123. |
| [29] | Neubauer S C, Givler K, Valentine S K, et al. Seasonal patterns and plant-mediated controls of subsurface wetland biogeochemistry[J]. Ecology, 2005, 86 (12): 3334-3344. |
| [30] | Dubinsky E A, Silver W L, Firestone M K. Tropical forest soil microbial communities couple iron and carbon biogeochemistry[J]. Ecology, 2010, 91 (9): 2604-2612. |
| [31] | Kappler A, Benz M, Schink B, et al. Electron shuttling via humic acids in microbial iron (Ⅲ) reduction in a freshwater sediment[J]. FEMS Microbiology Ecology, 2004, 47 (1): 85-92. |
| [32] | Weber K A, Urrutia M M, Churchill P F, et al. Anaerobic redox cycling of iron by freshwater sediment microorganisms[J]. Environmental Microbiology, 2006, 8 (1): 100-113. |
| [33] | Stumm W, Morgan J J. Aquatic chemistry: chemical equilibria and rates in natural waters[M]. New York: John Wiley and Sons, 2012. |
| [34] | Thamdrup B. Bacterial manganese and iron reduction in aquatic sediments[A]. In: Schink B (ed). Advances in Microbial Ecology, Vol. 16[C]. New York: Kluwer Academic/Plenum Publishers, 2000. 41-84. |
| [35] | 曲东, Schnell S. 纯培养条件下不同氧化铁的微生物还原能力[J]. 微生物学报, 2001, 41 (6): 745-749. |