 2014, Vol.
2014, Vol. 


青岛近海生物气溶胶中可培养微生物浓度及群落多样性的季节变化
引用本文
祁建华, 武丽婧, 高冬梅, 金川. 青岛近海生物气溶胶中可培养微生物浓度及群落多样性的季节变化[J]. 环境科学, 2014, (3): 801-809.
QI Jian-hua, WU Li-jing, GAO Dong-mei, JIN Chuan. Concentration and Community Diversity of Microbes in Bioaerosols in the Qingdao Coastal Region[J]. Environmental Science, 2014, (3): 801-809.
青岛近海生物气溶胶中可培养微生物浓度及群落多样性的季节变化
1. 中国海洋大学环境科学与工程学院, 青岛 266100;
2. 中国海洋大学海洋环境与生态教育部重点实验室, 青岛 266100
2. 中国海洋大学海洋环境与生态教育部重点实验室, 青岛 266100
摘要:为了研究近海生物气溶胶中可培养微生物浓度和群落多样性,于2009年7月~2010年6月在青岛两个采样点连续采集生物气溶胶样品,分析了其中陆源细菌、海源细菌、陆源真菌和海源真菌的浓度,并计算了Shannon-Weiner指数、Simpson's指数和Pielou指数. 结果表明,陆源细菌和海源细菌月均浓度分别为12~436 CFU·m-3和25~561 CFU·m-3,陆源真菌和海源真菌月均浓度分别为0~817 CFU·m-3和11~1346 CFU·m-3之间. 陆源细菌、海源细菌、陆源真菌和海源真菌浓度在冬季月份较低,2月达到最低值,在春夏月份较高. 海源微生物对总可培养类微生物的贡献高于陆源,平均占63%. 可培养微生物物种数在17~102之间,与微生物浓度具有一定的相关性,但并未呈现出明显的季节变化. 3种指数表明,生物气溶胶中陆源细菌、海源细菌、陆源真菌和海源真菌的群落结构在2月最简单,1月、11月和5月群落多样性较高,群落多样性与浓度的季节变化特征并不一致,而且不同类别的微生物群落存在季节和空间差异.
关键词:
生物气溶胶
群落多样性
细菌
真菌
青岛
Concentration and Community Diversity of Microbes in Bioaerosols in the Qingdao Coastal Region
1. College of Environmental Science and Engineering, Ocean University of China, Qingdao 266100, China;
2. Key Laboratory of Marine Environment and Ecology, Ministry of Education, Ocean University of China, Qingdao 266100, China
2. Key Laboratory of Marine Environment and Ecology, Ministry of Education, Ocean University of China, Qingdao 266100, China
Abstract: Bioaerosol samples were collected in Qingdao coastal region during July 2009 -June 2010 to investigate the concentration and community diversity of microbes in bioaerosols. Microbe concentrations (bacteria and fungi) in marine and terrestrial bioaerosols were determined and diversity indices including Shannon-Weiner index, Simpson's index and Pielou index were calculated in this study. Monthly average concentrations of terrestrial bacteria, marine bacteria, terrestrial fungi and marine fungi were in the ranges of 12-436 CFU·m-3, 25-561 CFU·m-3, 0-817 CFU·m-3 and 11-1346 CFU·m-3, respectively. There were consistent seasonal variations of these four types of microbe, with higher concentrations in spring and summer and lowest during winter, especially in February. Compared to terrestrial microbes, marine microbes account for higher proportion to the total culturable microbes, with a percentage of 63%. The number of microbial species varied from 17 to 102, and was partially correlated with microbial concentrations, however, it did not show obvious seasonal variation. Based on the analysis of calculated diversity indices, we found that the community diversities of four types of microbe were much higher in January, November and May than in February. The community diversity varied with the season, space and different microbial species, and showed a different seasonal variation from the microbial concentration.
Key words:
bioaerosols
community diversity
bacteria
fungi
Qingdao
生物气溶胶是大气气溶胶的重要组成部分,对空气质量和人体健康具有明显的影响[1, 2]. 生物气溶胶还可以改变大气中冰核(ice nuclei,IN)和云凝结核(cloud condensation nuclei,CCN)的数量和特性[3~8],进而影响云量和气候变化. 生物气溶胶主要由含有微生物或生物大分子等生命活性物质的生物微粒组成,包括细菌、 真菌、 病毒、 花粉、 孢子、 动植物碎裂分解体等具有生命活性的粒子[9, 10]. 而微生物是生物气溶胶中重要的组成部分,其微生物群落可以反映生物气溶胶中微生物的种类、 数量和组成特征,不同的群落结构对空气质量和人体健康的影响不同,同时微生物群落的变化也可以反映出环境因素对空气质量的影响. 因此,研究微生物群落多样性对于全面评价生物气溶胶对于人体健康、 环境空气质量及气候变化的影响具有重要的意义.
目前国内外对生物气溶胶中微生物群落特征和多样性所展开的研究中,Jeon等[11]在韩国观察到生物气溶胶中的细菌群落在亚洲沙尘事件中发生了突然改变. Maki等[12]调查了日本Suzu城生物气溶胶中耐盐细菌的群落结构,发现微生物群落在600 m和10 m高度相差较大,600 m高度的微生物群落虽然没有10 m的丰富,但是具有独特的微生物种类,可能源于长距离传输的沙尘. Lee等[13]分析了2007年中秋到冬末朝鲜半岛中部生物气溶胶中细菌和真菌的群落特征,研究发现颗粒物中细菌和真菌的多样性很高,包括潜在病原体和过敏原; 细菌和真菌群落的多样性存在季节变化. Bowers等[14]发现生物气溶胶在近地表层>0.5 μm的气溶胶粒子中占了很大的比例(平均22%),细菌群落组成具有明显的季节变化,这种变化主要原因是不同季节的局地陆源环境的差异,而不是季节更替的大气条件. Munday等[15]收集了不同高度(最高到150 m)的生物气溶胶样品,鉴定出15个细菌种群,细菌群落具有高多样性,而且群落多样性随高度而降低. 郑芷青等[16]对珠三角城市群气溶胶的研究指出,城市细菌的多样性趋势为:交通枢纽>工业区>商业区>居住区>绿化带. 夏晓敏等[17]发现厦门空气气溶胶中的细菌类群主要由厚壁菌门及变形细菌门的细菌构成,与其他城市相比厦门的空气细菌多样性较低. 已有的研究表明,不同地区的生物气溶胶微生物群落都具有一定的多样性,群落组成具有明显的空间变化,且群落多样性随采样高度和沙尘等特殊天气而变化,但目前关于生物气溶胶群落多样性的季节变化特征仍很缺乏,尤其是沿海生物气溶胶. 因此, 研究沿海地区生物气溶胶中微生物的群落多样性对于全面了解生物气溶胶的环境效应尤为重要.
本研究基于2009年7月~2010年6月在青岛地区连续采集的生物气溶胶样品,测定了样品中陆源和海源细菌及真菌的浓度,系统分析了青岛近海地区生物气溶胶中群落多样性的季节变化.
1 材料与方法
1.1 样品采集
在青岛地区设置了两个生物气溶胶采样点(图1),一个采样点位于中国海洋大学鱼山校区测试中心楼顶(36°03′N,120°19′E,距地面高度约为7.0 m),距海边1.0 km左右,附近为居民生活区,四周绿地面积约为50%; 另一个采样点位于中国海洋大学崂山校区教学楼六区楼顶(36°09′N,120°29′E,距地面高度约9.0 m),离海7 km,四周为学校教学区,绿地面积可达50%.
 | 图1 采样点示意 Fig.1 Location of the sampling site |
于2009年7月~2010年6月在两个采样点运用FA-1筛孔撞击式空气微生物采样器(辽阳康洁仪器研究所)采集可培养类微生物样品. 每月分别在上、 中、 下旬采样一次,每次间隔10 d左右; 采样时间为早上08:00. 在沙尘天气时加采样品. 采样器高度为1.5 m,采样流量为28.3 L ·min-1. 可培养陆源细菌和可培养海源细菌样品的采集时间为8 min,可培养陆源真菌和可培养海源真菌样品的采集时间为5 min; 总微生物样品的采集时间为30 min[18].
采用9.0 cm的玻璃平板进行生物气溶胶收集,平板灭菌后在无菌条件下加入20 mL的培养基. 陆源细菌、 陆源真菌、 海源细菌和海源真菌样品的培养基以及培养条件见表1[19]. 培养结束后分别对各级采样平板进行菌落计数以及形态观察.
| 表1 四类微生物的培养基以及培养条件 Table 1 Culture medium and conditions of four kinds of microbes |
1.2 浓度计算
运用Macher[20]提出的方法,对空气中细菌和真菌浓度进行了校正,并依据下式计算了生物气溶胶中微生物浓度:
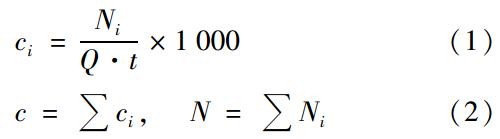
1.3 指数计算
香农多样性指数H′和辛普森指数D的计算公式如下:
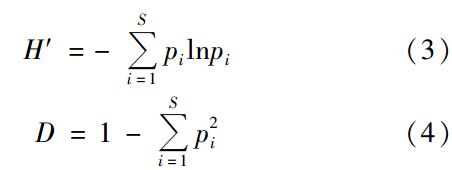
Pielou均一度指数用J表示:

2 结果与讨论
2.1 青岛近海生物气溶胶中可培养类微生物浓度月变化特征
2009年7月~2010年6月在青岛鱼山采样点和崂山采样点采集了可培养生物气溶胶样品,通过对每月上、 中、 下旬的样品进行体积加权平均,得到陆源细菌、 海源细菌、 陆源真菌和海源真菌的月平均浓度值.
2.1.1 青岛近海生物气溶胶中可培养细菌浓度的月际变化特征
图2(a)为2009年7月~2010年6月鱼山采样点陆源细菌及海源细菌的月平均浓度变化. 鱼山生物气溶胶中陆源细菌浓度与海源细菌的相关性系数为0.902,在P<0.01水平上显著相关. 鱼山采样点陆源和海源细菌月浓度变化范围分别为9~375 CFU ·m-3和25~532 CFU ·m-3. 陆源细菌及海源细菌月平均浓度最高值均出现在2009年12月,其他月份中5月较高,2月和6月浓度较低. 从图2(b)中可以看出,崂山陆源细菌的月均浓度最高值出现在2009年12月,为436 CFU ·m-3,随后9~10月出现较高值,最低值22 CFU ·m-3出现在6月,2~4月浓度也较低. 海源细菌5月浓度最高,为561 CFU ·m-3,12月浓度较高(474 CFU ·m-3),6月浓度最低,仅为40 CFU ·m-3. 尽管两个采样点陆源细菌和海源细菌的变化不尽相同,但是12月均出现了浓度高值,这是因为12月26日采集到沙尘样品,研究发现沙尘期间生物气溶胶浓度会显著增加[18, 21],所以引起12月浓度明显高于其它月份. 2010年2月温度相对较低(-2.5~3℃),不适于微生物的存活,所以细菌出现了最低值. 4月5日、 16日,6月5日、 16日样品采集时均有不同程度的雾,研究表明空气细菌浓度受雾滴的温度、 化学组成和酸碱度的影响较大[22],因此4月、 6月细菌浓度也较低. 5月平均气温为14~19℃,相对湿度为55%~86%,气候适宜微生物的生存,而且春季植被生长繁茂,微生物的来源较多,这是5月出现高浓度细菌的原因. 两个采样点陆源和海源细菌浓度的月际变化具有一定差异性,Adhikari等[23]的研究表明不同采样点对生物气溶胶的分布有着影响,可能是因为微生物来源和环境因素的不同.
 | 图2 青岛近海生物气溶胶中细菌浓度的月变化 Fig.2 Monthly variation of bacterial concentrations in the Qingdao coastal region |
鱼山和崂山采样点生物气溶胶中海源细菌占总细菌的平均百分比为60.1%和61.0%,海源细菌的贡献高于陆源细菌,可见青岛沿海区域的生物气溶胶组成明显受到海洋的影响.
2.1.2 青岛近海生物气溶胶中可培养真菌浓度的月际变化特征
图3(a)为2009年7月~2010年6月鱼山采样点陆源和海源真菌浓度的月变化,从中可以看出鱼山生物气溶胶中陆源真菌月平均浓度最高值出现在2010年5月,最低值出现在2009年10月和2010年3月,月平均浓度变化范围为0~516 CFU ·m-3. 而海源真菌月平均浓度最高值出现在2009年10月,最低值出现在2010年2月,月平均浓度变化范围为11~805 CFU ·m-3. 图3(b)为崂山采样点陆源真菌和海源真菌浓度的月变化特征. 崂山陆源真菌浓度的变化与鱼山类似,在2009年10月和2010年2月和3月的陆源真菌浓度最低,均为2 CFU ·m-3左右,从2010年4月起浓度开始升高,在6月到最高值,817 CFU ·m-3. 崂山生物采样点海源真菌浓度在100~1346 CFU ·m-3之间,月变化与鱼山采样点类似,浓度最高值出现在2009年10月,最低值出现在2010年2月,在9月、 5月浓度都较高.
2009年5月陆源和海源真菌的浓度都很高,此时气候适宜微生物的生存,而且春季植被生长繁茂,造成真菌浓度增加. 2009年10月两个采样点海源真菌浓度很高,与之前对青岛生物气溶胶的研究结果一致[19],采样期间盛行东南风,由海洋吹向陆地,这可能是海源真菌浓度很高而陆源真菌浓度很低的原因. 2009年8月、 9月海源和陆源真菌浓度相对较高,是因为当地夏季温度较高,平均为24℃,适于真菌粒子的生长. 已有的研究也发现,在一定的环境温度范围内,真菌更容易在较高温度下存活,而在低温时则受到抑制[18,24,25]. 因此真菌在夏季浓度较高,而在温度很低的2月出现了浓度的低值. 如前所述,6月因为出现了雾天,细菌浓度受到影响出现了较低值,而真菌浓度较高,这是因为真菌孢子对极端天气的耐受力要强于细菌[26]. 鱼山采样点2009年12月陆源真菌浓度高达494 CFU ·m-3,与细菌类似,浓度增大可能与12月26号采集到的沙尘样品有关. 但是崂山采样点陆源真菌浓度的增加远没有鱼山显著. 总体而言,崂山采样点陆源和海源真菌月平均浓度略高于鱼山. 除个别月份,崂山采样点生物气溶胶中真菌的月变化与鱼山采样点相近,这表明真菌浓度的分布具有一定的规律性,但是也存在空间分布差异,表明真菌浓度受到不同地区来源和环境因素的影响.
鱼山和崂山采样点生物气溶胶中海源真菌占总真菌的平均百分比为59.1%和71.2%,海源真菌的贡献高于陆源细菌,可见海洋对沿海区域的生物气溶胶组成有明显的影响,与已有的研究结果一致[19, 27].
 | 图3 青岛近海生物气溶胶中真菌浓度的月变化 Fig.3 Monthly variation of fungi concentrations in the Qingdao coastal region |
2.2 青岛近海生物气溶胶中可培养微生物群落的月季特征
2.2.1 青岛近海生物气溶胶总可培养微生物群落的月季分布
2009年11月~2010年6月青岛近海生物气溶胶中总可培养微生物浓度(细菌和真菌浓度之和)及物种数随月份变化较大. 鱼山采样点2010年2月可培养微生物浓度最低,为73 CFU ·m-3,微生物物种数目最少,为17. 5月可培养微生物浓度最高,达1948 CFU ·m-3,微生物物种数最高,为102. 崂山采样点的微生物物种数变化与鱼山物类似,2月最低,6月最高,变化范围为39~90. 2009年12月鱼山和崂山采样点的微生物浓度很高,相应的物种数也很高,分别为102和88,比11月增加59%和57%,可见沙尘天气不仅对微生物浓度影响显著,而且对微生物物种数目的增加也起到显著作用,沙尘期间应该有外来微生物长距离输入研究区域. Jeon等[11]在韩国观察到生物气溶胶中的细菌群落在亚洲沙尘事件中发生了突然改变. 研究发现,沙尘传输过几百乃至几千公里后,耐盐菌成为生物气溶胶中微生物的典型种类[28, 29]. 可见,沙尘天气对生物气溶胶中微生物群落结构及优势微生物的影响非常大. 除个别月份外,两个采样点可培养微生物浓度和物种数目的变化趋势类似. 鱼山和崂山采样点可培养微生物月平均浓度和物种数目的相关系数分别为0.835和0.827,均在P<0.01水平上显著相关.
 | 图4 青岛近海生物气溶胶中可培养微生物多样性指数的月际变化Fig.4 Monthly variation of the diversity index of total culturable microbes in the Qingdao coastal region |
从季节上看,鱼山采样点可培养微生物浓度的季节分布为春季>秋季>冬季>夏季,而崂山采样点与鱼山不同,可培养微生物浓度秋季与春季接近,略高于夏季,而冬季最低,这是因为夏季气温高、 紫外线强,而秋季气候温和、 适宜微生物的生长繁殖. 两个采样点可培养微生物浓度都是秋季较高,与之前对青岛[30]、 广州[31]和北京[32]的研究结果一致. Adhikari 等[33]也在美国俄亥俄州观察到真菌浓度的最高值出现在秋季. 另外一些研究发现生物气溶胶中可培养微生物浓度在夏季出现浓度最大值,而冬季和春季浓度较低[34~36],这是因为这些地区夏季温度较高适宜微生物生长所致. 可见,生物气溶胶中可培养微生物浓度的季节分布特征因不同地区的气象条件不同而存在显著的空间差别.
鱼山采样点可培养微生物浓度春季为855 CFU ·m-3,明显高于冬季的741 CFU ·m-3,但物种数相差不大,春、 冬季分别为67和64. 与鱼山类似,崂山可培养微生物浓度也是春季高于冬季,浓度分别为1057 CFU ·m-3和703 CFU ·m-3,物种数也是春季(72)高于冬季(67). 由此可见,与微生物浓度存在显著的季节变化不同,物种数的季节变化并不明显. 崂山采样点季节平均的微生物物种数目略高于鱼山采样点.
图4是2009年11月~2010年6月青岛近海生物气溶胶中可培养微生物的3种多样性指数的月际分布. Shannon-Weiner指数通常多用于反映群落的复杂程度,指数越大,则群落越复杂,对环境的反馈功能越强,从而使群落得到较大的缓冲,更趋于稳定. Simpson's指数即优势度指数,指数越大群落多样性越高. Pielou指数即均匀度指数,反映了各物种个体数目分配的均匀程度,指数越大,物种数目分配越均匀. 由图4(a)可以看出,Shannon-Weiner指数2010年1月、 2月和6月崂山采样点的Shannon-Weiner指数大于鱼山,其他月份则是鱼山采样点的指数更大,而且鱼山采样点的物种数高于崂山,说明该区域的微生物群落相对复杂. 鱼山和崂山采样点可培养微生物的Shannon-Weiner指数分别在2.68~3.99和3.14~3.89之间,月际分布并不完全相同. 两个采样点Shannon-Weiner指数最低值都出现于2012年2月,鱼山采样点的Shannon-Weiner指数5月最高,11月次之,而崂山采样点则是1月最高,11月和6月次之.
2010年2月两个采样点的微生物浓度和物种数目最低,所以Shannon-Weiner指数也是最低. 11月份鱼山和崂山的物种数分别是64和56,但是Simpson's指数和Pielou指数都很高,说明物种数目分布均匀、 优势菌较少,所以群落多样性高,相应地Shannon-Weiner指数也较高. 2009年12月和2010年5月,崂山采样点的微生物浓度和物种数目都属于较高水平,但是其Shannon-Weiner指数并不是最高. 从图4(b)和4(c)可以看出12月的Simpson's指数和Pielou指数较低,说明该月份优势菌较多而且各物种数目分配的不均匀,所以微生物浓度虽然高,但是群落反而相对简单,多样性不高. 鱼山采样点5月份物种数高达102,远高于崂山(85),而且鱼山的Simpson's指数和Pielou指数都明显高于崂山,说明鱼山物种数比崂山分布均匀而且群落多样性高,所以鱼山Shannon-Weiner指数在5月出现了最大值但崂山Shannon-Weiner指数较低. 2010年1月两个采样点的微生物浓度并不高,但是3种指数都很高,而且物种数目相对较高,说明物种分布相对均匀,优势菌相对较少,群落多样性高.
鱼山和崂山采样点的Pielou指数和微生物浓度的相关性系数分别为-0.773和-0.858,在P<0.05和P<0.01水平上呈显著负相关,这说明微生物浓度大的月份往往存在优势度较大的菌种,导致Pielou指数降低,如2009年12月和2010年5月两采样点的样品; 而2009年11月和2010年2月微生物浓度较小,Pielou指数较高,说明物种分配比较均匀. 除了2010年1月,鱼山采样点的Pielou指数均高于崂山采样点,这说明鱼山采样点微生物群落中物种分配比崂山相对均匀.
除个别月,崂山采样点可培养微生物浓度高于鱼山,但是微生物物种数、 Shannon-Weiner指数和Simpson's指数低于鱼山,这表明崂山采样点优势菌对微生物浓度的贡献高于鱼山,而鱼山采样点的群落比崂山采样点多样性更高,群落更复杂、 稳定.
2.2.2 青岛近海生物气溶胶中不同类别微生物的群落的月际变化
从表2可知,2009年11月~2010年6月,陆源细菌、 海源细菌、 陆源真菌和海源真菌的物种数随着月季变化很大. 由表2可知,青岛近海生物气溶胶中不同来源可培养微生物浓度和物种数基本上呈显著正相关(P<0.05),其中崂山采样点陆源真菌浓度和物种数的相关性在P<0.01程度上呈显著正相关; 鱼山采样点海源细菌浓度和物种数并不相关. 这说明当不同季节,生物气溶胶来源和环境条件不同,引起生物气溶胶中微生物物种增加或减少的时候,通常会引起微生物浓度升高或降低.
两个采样点陆源细菌的物种数略高于海洋细菌(表3),其Shannon-Weiner指数、 Simpson's指数和Pielou指数均略高于海洋细菌,与海源细菌相比,陆源细菌的微生物群落中物种数分布均匀,优势菌相对较少,生物多样性高,群落较复杂. 崂山采样点的陆源、 海源物种数和生物多样性略高于鱼山. 与总可培养微生物类似,鱼山样点陆源和海源细菌的Shannon-Weiner指数最低值都出现于2月,这是因为2月温度低,不利于微生物生长,来源少,微生物浓度和物种数目最低. 而崂山采样点的情况较为复杂,陆源细菌的Shannon-Weiner指数在6月出现最低值,而海源细菌在12月出现了最低值. 这是因为6月陆源细菌的物种数最低,只有11,而且Simpson's指数较低,说明微生物种类少,而且存在优势菌,所以6月微生物群落多样性很低. 12月海源细菌虽然浓度高但物种数为19,Simpson's指数和Pielou指数分别为0.76和0.68,远低于其它月份,说明微生物种类较少,而且物种数分配不均匀,优势菌相对较多,因此微生物群落相对简单,多样性低. 鱼山采样点陆源和海源细菌Shannon-Weiner指数的最高值分别出现在12月和11月,而物种数分别为16和30,崂山采样点陆源和海源细菌Shannon-Weiner指数的最高值出现在1月和2月. 陆源和海源细菌在相应月份的物种数较高,而且Simpson's指数和Pielou指数都很高,表明微生物物种数分布均匀,优势菌相对较少,群落相对复杂,生物多样性高. 可见,陆源和海源细菌的群落特征和变化并不完全相同,而且两个采样点也因为来源和环境条件的不同而存在空间差异.
除了鱼山冬季外,两个采样点海源真菌的物种数和Shannon-Weiner指数均为略高于陆源真菌,这表明海源真菌的群落结构比陆源真菌较为复杂、 多样性更高. 两个采样点陆源真菌和海源真菌的物种数随季节变化较大,没有明显的贡献优势. 两个采样点陆源真菌的Shannon-Weiner指数都是在12月最高,这是因为12月陆源真菌物种数很高,Simpson's指数和Pielou指数也很高,表明微生物物种分布均匀,多样性高. 鱼山和崂山采样点陆源真菌Shannon-Weiner指数的最低值分别出现在3月和2月,因为物种数很低,分别为3和2,所以微生物群落多样性很低. 鱼山和崂山海源真菌Shannon-Weiner指数的最高值分别出现在5月和6月,这是因为相应月份物种丰富,物种数高达33和30,而且Simpson's指数和Pielou指数也很高,表面物种数分布均匀,微生物群落比较复杂. 鱼山和崂山海源真菌Shannon-Weiner指数的最低值均出现在2月,如前所述,是因为2月物种数非常低,因此微生物群落简单. 同样,陆源和海源细菌的群落特征和变化并不完全相同,而且两个采样点也存在空间差异.
| 表2 青岛近海生物气溶胶中不同来源可培养微生物的浓度和物种数目相关性 Table 2 Correlation of different kinds of culturable microbial concentration and species number in the Qingdao coastal region |
| 表3 采样点各类微生物的物种数和3种多样性指数Table 3 Species number and three diversity indexes of different kinds of microbes in the sampling sites |
总体而言,不同地区、 不同季节、 不同类别的微生物群落结构存在差异,主要是受到微生物来源和环境条件的影响,微生物浓度并不能反映微生物群落的多样性.
3 结论
(1)生物气溶胶中陆源细菌、 海源细菌、 陆源真菌和海源真菌浓度在冬季月份较低,在春夏月份较高,可培养微生物浓度的最低值均出现在温度较低的2月. 不同种类的细菌和真菌月季变化并不完全一致,这是来源和不同物种对环境条件耐受力的不同所致.
(2)沙尘天气下微生物浓度明显升高,而雾天会使细菌浓度降低.
(3)海源微生物对总可培养微生物的贡献高于陆源,可见海洋对沿海区域的生物气溶胶组成有明显的影响.
(4)生物气溶胶中微生物物种数目和微生物浓度具有一定的相关性,微生物浓度越高,物种数目也越多. 微生物浓度存在显著的季节变化,但物种数的季节变化并不明显.
(5)生物气溶胶中陆源细菌、 海源细菌、 陆源真菌和海源真菌的群落结构在2月最简单,1月、 11月和5月群落多样性较高,而群落多样性与浓度的分布特征并不一致,可见,微生物浓度并不能反映微生物群落的多样性. 要全面评价微生物对空气质量和人体健康的影响,不仅需要研究浓度分布,更需要对群落结构深入研究.
(6)不同类别的生物气溶胶中微生物群落多样性存在季节和空间差异,主要是受到微生物来源、 环境条件以及为微生物物种特性的影响.
参考文献
| [1] | Kalogerakis N, Paschali D, Lekaditis V, et al. Indoor air quality—bioaerosol measurements in domestic and office premises[J]. Journal of Aerosol Science, 2005, 36 (5-6): 751-761. |
| [2] | Ho J, Duncan S. Estimating aerosol hazards from an anthrax letter[J]. Journal of Aerosol Science, 2005, 36 (5-6): 701-719. |
| [3] | Vali G, Christensen M, Fresh R, et al. Biogenic ice nuclei. Part II: Bacterial sources[J]. Journal of Atmospheric Sciences, 1976, 33 (8): 1565-1570. |
| [4] | Vaïtilingom M, Attard E, Gaiani N, et al. Long-term features of cloud microbiology at the puy de D?me (France)[J]. Atmospheric Environment, 2012, 56: 88-100. |
| [5] | Maki L R, Galyan E L, Chang-Chien M M, et al. Ice nucleation induced by Pseudomonas syringae[J]. Applied Microbiology, 1974, 28 (3): 456-459. |
| [6] | Garcia E, Hill T C, Prenni A J, et al. Biogenic ice nuclei in boundary layer air over two U.S. High Plains agricultural regions[J]. Journal of Geophysical Research, 2012, 117 (D18), doi: 10.1029/2012JD018343. |
| [7] | Franc G D, DeMott P J. Cloud activation characteristics of airborne Erwinia carotovora cells[J]. Journal of Applied Meteorology, 1998, 37 (10): 1293-1300. |
| [8] | Bauer H, Giebl H, Hitzenberger R, et al. Airborne bacteria as cloud condensation nuclei[J]. Journal of Geophysical Research, 2003, 108 (D21), doi: 10.1029/2003JD003545. |
| [9] | 胡家骏, 周群英. 环境工程微生物学[M]. 北京: 高等教育出版社, 1998. |
| [10] | Mandrioli P. Basic aerobiology[J]. Aerobiologia, 1998, 14 (2-3): 89-94. |
| [11] | Jeon E M, Kim H J, Jung K, et al. Impact of Asian dust events on airborne bacterial community assessed by molecular analyses[J]. Atmospheric Environment, 2011, 45 (25): 4313-4321. |
| [12] | Maki T, Susuki S, Kobayashi F, et al. Phylogenetic analysis of atmospheric halotolerant bacterial communities at high altitude in an Asian dust (KOSA) arrival region, Suzu City[J]. Science of the Total Environment, 2010, 408 (20): 4556-4562. |
| [13] | Lee S H, Lee H J, Kim S J, et al. Identification of airborne bacterial and fungal community structures in an urban area by T-RFLP analysis and quantitative real-time PCR[J]. Science of the Total Environment, 2010, 408 (6): 1349-1357. |
| [14] | Bowers R M, McCubbin I B, Hallar A G, et al. Seasonal variability in airborne bacterial communities at a high-elevation site[J]. Atmospheric Environment, 2012, 50: 41-49. |
| [15] | Munday C I, O'Loingsigh T, Tapper N J, et al. Utilisation of Rep-PCR to track microbes in aerosols collected adjacent to their source, a saline lake in Victoria, Australia[J]. Science of the Total Environment, 2013, 450-451: 317-325. |
| [16] | 郑芷青, 谢小保, 欧阳友生, 等. 珠江三角洲城市群大气微生物优势种群及时空分异特征[J]. 地理研究, 2009, 28 (3): 625-633. |
| [17] | 夏晓敏, 汪建君, 陈立奇, 等. 厦门市区10月份大气气溶胶中细菌群落结构的初步研究[J]. 厦门大学学报(自然科学版), 2010, 49 (5): 682-687. |
| [18] | Li M F, Qi J H, Zhang H D, et al. Concentration and size distribution of bioaerosols in an outdoor environment in the Qingdao coastal region[J]. Science of the Total Environment, 2011, 409 (19): 3812-3819. |
| [19] | 刘苗苗, 祁建华, 高冬梅, 等. 青岛近海秋季生物气溶胶分布特征[J]. 生态环境, 2008, 17 (2): 565-571. |
| [20] | Macher J M. Positive-hole correction of multiple-jet impactors for collecting viable microorganisms[J]. American Industrial Hygiene Association Journal, 1989, 50 (11): 561-568. |
| [21] | Griffin D W, Kubilay N, Ko?ak M, et al. Airborne desert dust and aeromicrobiology over the Turkish Mediterranean coastline[J]. Atmospheric Environment, 2007, 41 (19): 4050-4062. |
| [22] | Fuzzi S, Mandrioli P, Perfetto A. Fog droplets-an atmospheric source of secondary biological aerosol particles[J]. Atmospheric Environment, 1997, 31 (2): 287-290. |
| [23] | Adhikari A, Sen M M, Gupta-Bhattacharya S, et al. Airborne viable, non-viable, and allergenic fungi in a rural agricultural area of India: a 2-year study at five outdoor sampling stations[J]. Science of the Total Environment, 2004, 326 (1): 123-141. |
| [24] | Fang Z, Ouyang Z, Hu L, Wang X, et al. Progresses of airborne microbial communities in urban ecosystem[J]. Aata Ecologica Sinica, 2004, 24 (2): 315-322. |
| [25] | Mitchell R, Gu J D. Environmental microbiology[M]. (2nd ed.). Beijing: Science Press, 2010. |
| [26] | Tang J W. The effect of environmental parameters on the survival of airborne infectious agents[J]. Journal of the Royal Society Interface, 2009, 6 (Suppl 6): S737-S746. |
| [27] | 陈皓文. 青岛空气微生物状况的测定[J]. 山东科学, 2003, 16 (1): 9-13. |
| [28] | Maki T, Susuki S, Kobayashi F, et al. Phylogenetic diversity and vertical distribution of a halobacterial community in the atmosphere of an Asian dust (KOSA) source region, Dunhuang city[J]. Air Quality, Atmosphere & Health, 2008, 1 (2): 81-89. |
| [29] | Kellogg C A, Griffin D W. Aerobiology and the global transport of desert dust[J]. Trends in Ecology & Evolution, 2006, 21 (11): 638-644. |
| [30] | 徐文兵, 祁建华, 金川, 等. 青岛近海夏、秋季生物气溶胶分布特征研究[J]. 环境科学, 2011, 32 (1): 9-17. |
| [31] | 欧阳友生, 谢小保, 陈仪本, 等. 广州市空气微生物含量及其变化规律研究[J]. 微生物学通报, 2006, 33 (3): 47-51. |
| [32] | Fang Z, Ouyang Z, Zheng H, et al. Culturable airborne bacteria in outdoor environments in Beijing, China[J]. Microbial Ecology, 2007, 54 (3): 487-496. |
| [33] | Adhikari A, Reponen T, Grinshpun S A, et al. Correlation of ambient inhalable bioaerosols with particulate matter and ozone: a two-year study[J]. Environmental Pollution, 2006, 140 (1): 16-28. |
| [34] | O'Gorman C M, Fuller H T. Prevalence of culturable airborne spores of selected allergenic and pathogenic fungi in outdoor air[J]. Atmospheric Environment, 2008, 42 (18): 4355-4368. |
| [35] | Matthias-Maser S, Reichert K, Jaenicke R. Primary biological aerosol particles at the high alpine site of Jungfraujoch/Switzerland[J]. Journal of Aerosol Science, 2000, 31: 955-956. |
| [36] | Lighthart B, Shaffer B T. Viable bacterial aerosol particle size distributions in the midsummer atmosphere at an isolated location in the high desert chaparral[J]. Aerobiologia, 1995, 11 (1): 19-25. |